

doi: 10.1038/nsmb.2301.
Translation drives mRNA quality control
Affiliations
- PMID: 22664987
- PMCID: PMC4299859
- DOI: 10.1038/nsmb.2301
Item in Clipboard
Translation drives mRNA quality control
Nat Struct Mol Biol.
.
Abstract
There are three predominant forms of co-translational mRNA surveillance: nonsense-mediated decay (NMD), no-go decay (NGD) and nonstop decay (NSD). Although discussion of these pathways often focuses on mRNA fate, there is growing consensus that there are other important outcomes of these processes that must be simultaneously considered. Here, we seek to highlight similarities between NMD, NGD and NSD and their probable origins on the ribosome during translation.
Figures
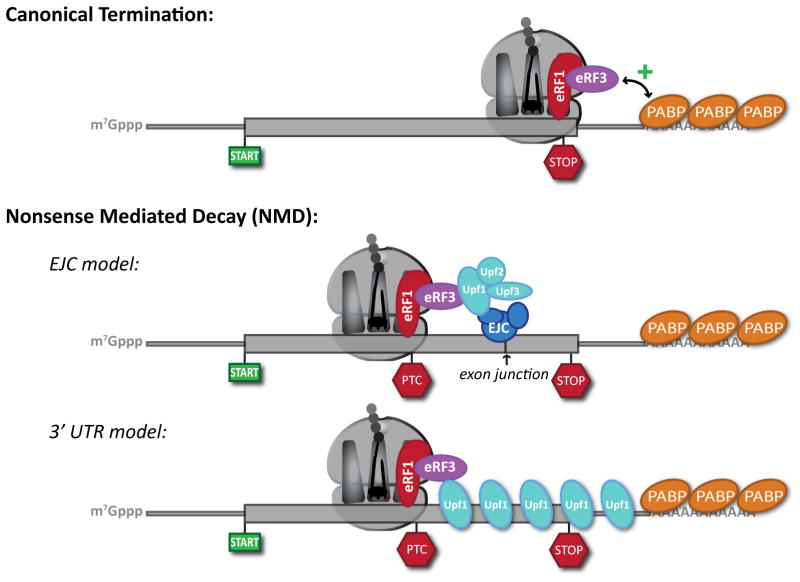
(a) Canonical termination. Capped and polyadenylated messages are translated through the open reading frame until recognition of a stop codon by the eukaryotic release factors, eRF1 and eRF3. Close proximity of authentic stop codons with the poly(A) tail is proposed to facilitate interactions between eRF3 and poly(A)-binding proteins (PABP) that positively contribute to peptide release. (b) Nonsense-mediated decay (NMD). In the case of a premature stop codon (PTC), lack of proximity is proposed to disrupt interaction between eRF3 and PABP. Canonical termination is further modified by the presence of NMD factors. In the EJC model of higher eukaryotes, this results from encountering a stop codon upstream of an exon-junction complex (EJC). In this model, communication between the termination factors and the EJC is effectively bridged by Upf1 in coordination with Upf2 and Upf3. In the 3′ UTR model, a PTC effectively extends the de facto 3′ untranslated region (UTR) of the message. This provides a larger binding platform for Upf1, which drives the termination event towards NMD rather than classical termination. Thick line, open reading frame; thin line, 5′ and 3′ UTR.
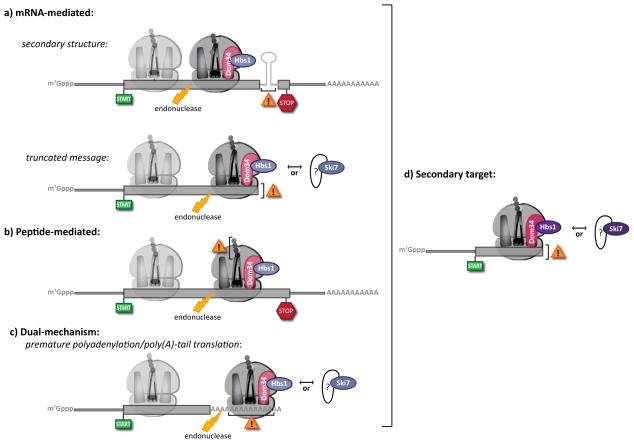
No-go decay (NGD) and non-stop decay (NSD) both involve the recognition of stalled ribosome complexes. These stalls can arise through multiple mechanisms. (a) mRNA-mediated targets. Inhibitory mRNA secondary structures stall ribosomes at internal loci (top), while truncated mRNAs result in terminal stalls (bottom). While classically distinguished as NGD and NSD targets, respectively, increasing evidence suggests that such distinctions belie common mechanistic features. (b) Peptide-mediated targets. Inhibitory peptide sequences lead to internally stalled ribosomes, classically defined as NGD substrates. (c) Dual-mechanism targets. Translation of the poly(A) tail, originally considered to mimic a truncated message and invoke NSD, likely induces ribosome stalling prior to its arrival at the end of the message. As such, the distinction between NGD and NSD under these conditions is ambiguous in the absence of further experimentation. In all cases, (a), (b) and (c), endonucleolytic cleavage occurs upstream of the stalled ribosome, potentially stimulated by Dom34 and Hbs1. This tentative role for Dom34 and Hbs1 prior to cleavage is indicated by the increased transparency of these factors in (a–c). Following cleavage, the trailing ribosome (shown transparently) elongates to the point of cleavage, generating an ideal target for Dom34/Hbs1 (or Ski7) recognition (d). At present, no Dom34-like factor has been identified that interacts with Ski7.
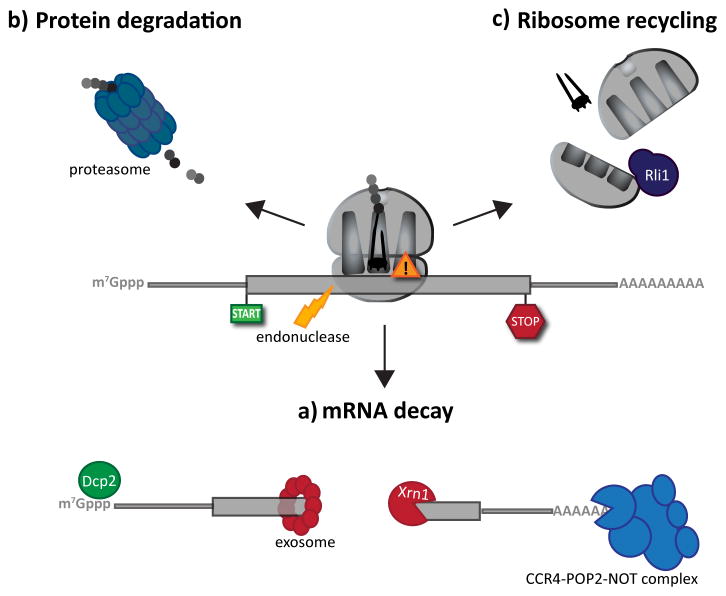
Following the recognition of NMD, NGD or NSD ribosome complexes, at least three discrete salvage pathways are invoked: mRNA decay, protein degradation and ribosome recycling. (a) mRNA decay. Endonucleolytic cleavage subverts the need for deadenylation, by the CCR4-POP2-NOT complex, and decapping, by Dcp2, prior to mRNA decay. Rapid mRNA degradation then proceeds through canonical means, including 5′-3′ degradation by Xrn1 and 3′-5′ degradation by the exosome. (b) Protein degradation. Targeted degradation of aberrant peptides occurs via the ubiquitin-proteasome system. Several E3 ligases have been implicated in this process, but the molecular features of substrate recognition remain to be determined. (c) Ribosome recycling. Dom34:Hbs1 are known to exploit the canonical recycling activities of Rli1 to effect ribosome recycling during NGD and NSD. Recycling of ribosome complexes during NMD are less well-characterized, but may involve Upf1.
References
-
- Isken O, Maquat LE. Quality control of eukaryotic mRNA: safeguarding cells from abnormal mRNA function. Genes & development. 2007;21:1833–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
