

Common features at the start of the neurodegeneration cascade
- PMID: 22666178
- PMCID: PMC3362641
- DOI: 10.1371/journal.pbio.1001335
Common features at the start of the neurodegeneration cascade
Abstract
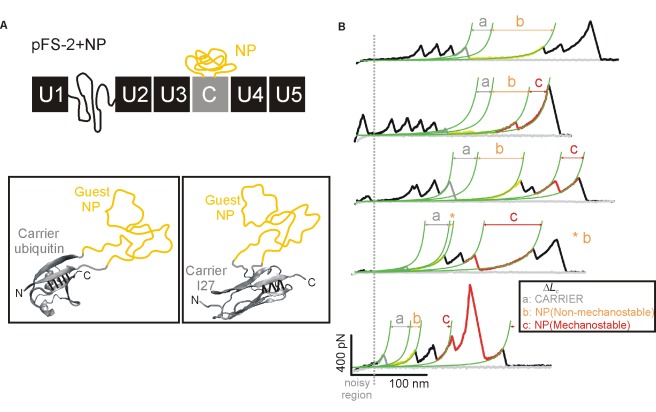
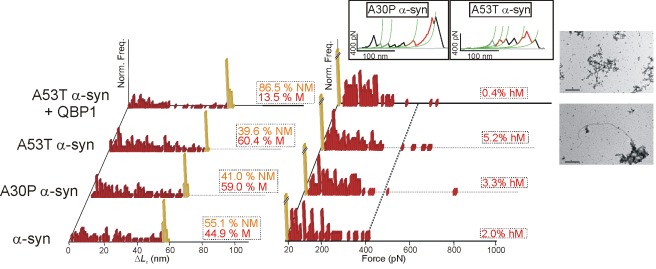
Amyloidogenic neurodegenerative diseases are incurable conditions with high social impact that are typically caused by specific, largely disordered proteins. However, the underlying molecular mechanism remains elusive to established techniques. A favored hypothesis postulates that a critical conformational change in the monomer (an ideal therapeutic target) in these "neurotoxic proteins" triggers the pathogenic cascade. We use force spectroscopy and a novel methodology for unequivocal single-molecule identification to demonstrate a rich conformational polymorphism in the monomer of four representative neurotoxic proteins. This polymorphism strongly correlates with amyloidogenesis and neurotoxicity: it is absent in a fibrillization-incompetent mutant, favored by familial-disease mutations and diminished by a surprisingly promiscuous inhibitor of the critical monomeric β-conformational change, neurotoxicity, and neurodegeneration. Hence, we postulate that specific mechanostable conformers are the cause of these diseases, representing important new early-diagnostic and therapeutic targets. The demonstrated ability to inhibit the conformational heterogeneity of these proteins by a single pharmacological agent reveals common features in the monomer and suggests a common pathway to diagnose, prevent, halt, or reverse multiple neurodegenerative diseases.
Conflict of interest statement
The authors declare a competing financial interest: M.C.-V., J.O., and R.H. are co-inventors on an international patent application (No. P 201031846,PCT/ES2011/070867) covering the results contained in this article. Any potential income generated by exploitation of the patent rights and received by their employer, the CSIC, shall be shared with these authors according to Spanish law and the regulations of the CSIC.
Figures



Comment in
-
Single-molecule technique links structural fluctuations of proteins to brain diseases.PLoS Biol. 2012;10(5):e1001338. doi: 10.1371/journal.pbio.1001338. Epub 2012 May 29. PLoS Biol. 2012. PMID: 22666179 Free PMC article. No abstract available.
Similar articles
-
Single-molecule technique links structural fluctuations of proteins to brain diseases.PLoS Biol. 2012;10(5):e1001338. doi: 10.1371/journal.pbio.1001338. Epub 2012 May 29. PLoS Biol. 2012. PMID: 22666179 Free PMC article. No abstract available.
-
[Modification of [PSI+] prion properties by the combination of amino acid changes within Sup35 protein N-domain].Mol Biol (Mosk). 2014 Mar-Apr;48(2):314-21. Mol Biol (Mosk). 2014. PMID: 25850301 Russian.
-
Unequivocal single-molecule force spectroscopy of intrinsically disordered proteins.Methods Mol Biol. 2012;896:71-87. doi: 10.1007/978-1-4614-3704-8_5. Methods Mol Biol. 2012. PMID: 22821518
-
Targeting the chameleon: a focused look at α-synuclein and its roles in neurodegeneration.Mol Neurobiol. 2013 Apr;47(2):446-59. doi: 10.1007/s12035-012-8334-1. Epub 2012 Sep 1. Mol Neurobiol. 2013. PMID: 22940885 Review. No abstract available.
-
Probing the role of structural features of mouse PrP in yeast by expression as Sup35-PrP fusions.Prion. 2012 Jul 1;6(3):201-10. doi: 10.4161/pri.19214. Epub 2012 Jul 1. Prion. 2012. PMID: 22449853 Free PMC article. Review.
Cited by
-
Nanotechnology-Based Drug Delivery Strategies to Repair the Mitochondrial Function in Neuroinflammatory and Neurodegenerative Diseases.Pharmaceutics. 2021 Dec 1;13(12):2055. doi: 10.3390/pharmaceutics13122055. Pharmaceutics. 2021. PMID: 34959337 Free PMC article. Review.
-
Direct Detection of α-Synuclein Dimerization Dynamics: Single-Molecule Fluorescence Analysis.Biophys J. 2015 Apr 21;108(8):2038-47. doi: 10.1016/j.bpj.2015.03.010. Biophys J. 2015. PMID: 25902443 Free PMC article.
-
Depicting Conformational Ensembles of α-Synuclein by Single Molecule Force Spectroscopy and Native Mass Spectroscopy.Int J Mol Sci. 2019 Oct 19;20(20):5181. doi: 10.3390/ijms20205181. Int J Mol Sci. 2019. PMID: 31635031 Free PMC article.
-
Neurodegenerative Diseases: Regenerative Mechanisms and Novel Therapeutic Approaches.Brain Sci. 2018 Sep 15;8(9):177. doi: 10.3390/brainsci8090177. Brain Sci. 2018. PMID: 30223579 Free PMC article. Review.
-
Protein aggregation and therapeutic strategies in SOD1- and TDP-43- linked ALS.Front Mol Biosci. 2024 May 24;11:1383453. doi: 10.3389/fmolb.2024.1383453. eCollection 2024. Front Mol Biosci. 2024. PMID: 38855322 Free PMC article. Review.
References
-
- Chiti F, Dobson C. M. Protein misfolding, functional amyloid and human disease. Ann Rev Biochem. 2006;75:333–366. - PubMed
-
- Uversky V. Intrinsic disorder in proteins associated with neurodegenerative diseases. In: Ovádi J, Orosz F, editors. Protein folding and misfolding: neurodegenerative diseases. New York: Springer-Verlag; 2009. pp. 21–75.
-
- Kayed R, Head E, Thompson J. L, McIntire T. M, Milton S. C, et al. Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science. 2003;300:486–489. - PubMed
-
- Nagai Y, Popiel H. A. Conformational changes and aggregation of expanded polyglutamine proteins as therapeutic targets of the polyglutamine diseases: exposed β-sheet hypothesis. Curr Pharm Des. 2008;14:3267–3279. - PubMed
-
- Nagai Y, Inui T, Popiel H. A, Fujikake N, Hasegawa K, et al. A toxic monomeric conformer of the polyglutamine protein. Nat Struct Biol. 2007;14:332–340. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
