

Bipartite tetracysteine display reveals allosteric control of ligand-specific EGFR activation
- PMID: 22667988
- PMCID: PMC3789377
- DOI: 10.1021/cb300216f
Bipartite tetracysteine display reveals allosteric control of ligand-specific EGFR activation
Abstract
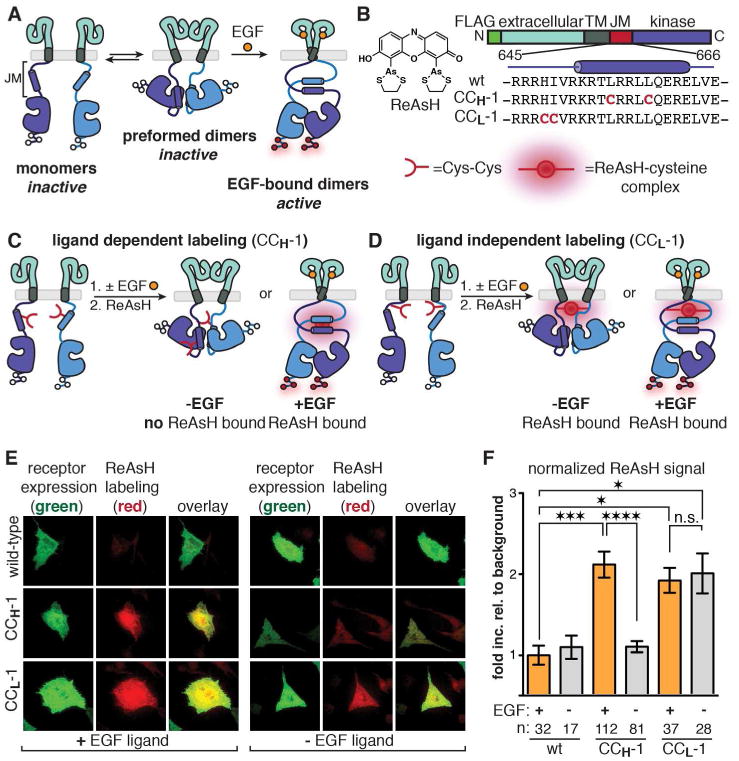
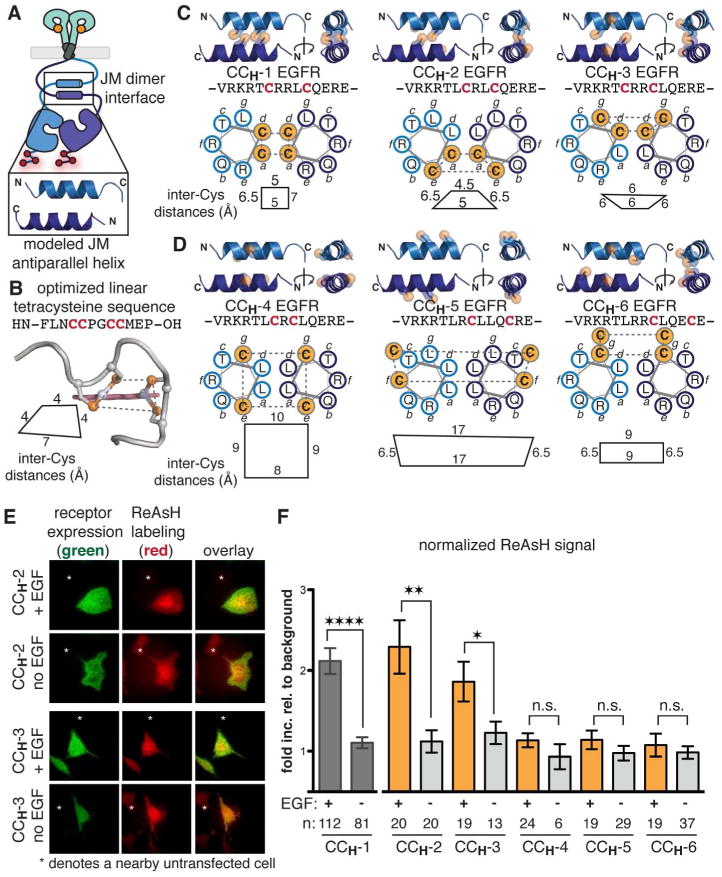
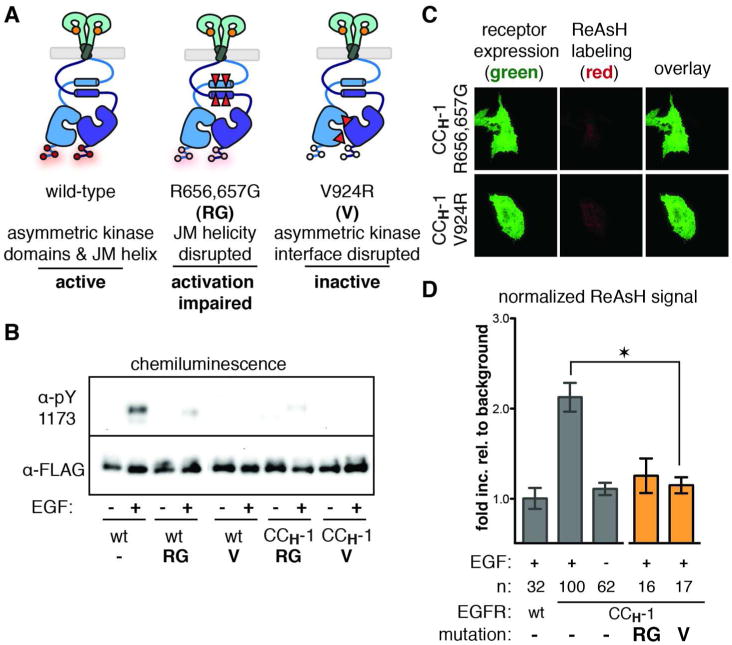
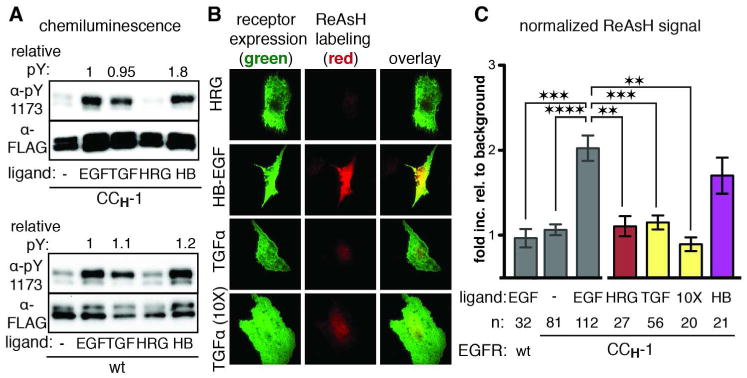
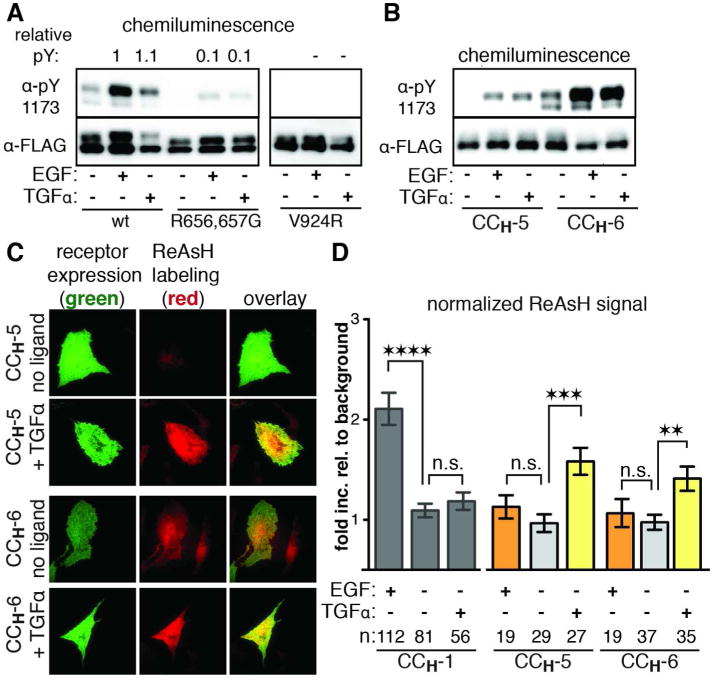
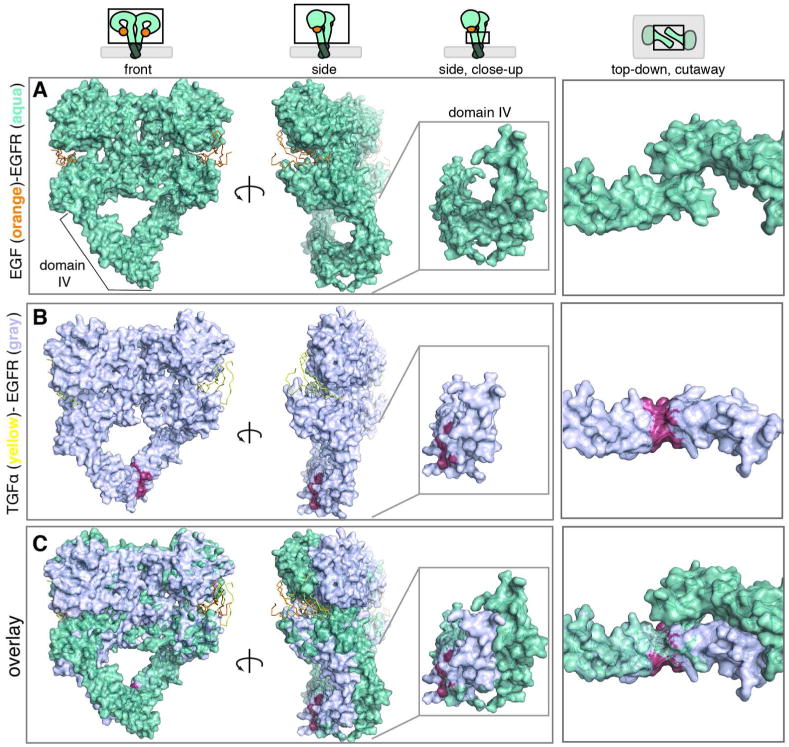
Aberrant activation of the epidermal growth factor receptor (EGFR), a prototypic receptor tyrosine kinase, is critical to the biology of many common cancers. The molecular events that define how EGFR transmits an extracellular ligand binding event through the membrane are not understood. Here we use a chemical tool, bipartite tetracysteine display, to report on ligand-specific conformational changes that link ligand binding and kinase activation for full-length EGFR on the mammalian cell surface. We discover that EGF binding is communicated to the cytosol through formation of an antiparallel coiled coil within the intracellular juxtamembrane (JM) domain. This conformational transition is functionally coupled to receptor activation by EGF. In contrast, TGFα binding is communicated to the cytosol through formation of a discrete, alternative helical interface. These findings suggest that the JM region can differentially decode extracellular signals and transmit them to the cell interior. Our results provide new insight into how EGFR communicates ligand-specific information across the membrane.
Figures
References
-
- Avraham R, Yarden Y. Feedback regulation of EGFR signalling: decision making by early and delayed loops. Nature Reviews Molecular Cell Biology. 2011;12(2):104–117. - PubMed
-
- Tao RH, Maruyama IN. All EGF(ErbB) receptors have preformed homo- and heterodimeric structures in living cells. Journal Of Cell Science. 2008;121(19):3207–3217. - PubMed
-
- Zhang X, Gureasko J, Shen K, Cole PA, Kuriyan J. An allosteric mechanism for activation of the kinase domain of epidermal growth factor receptor. Cell. 2006;125(6):1137–1149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
