

Matrix rigidity controls endothelial differentiation and morphogenesis of cardiac precursors
- PMID: 22669846
- PMCID: PMC11055637
- DOI: 10.1126/scisignal.2003002
Matrix rigidity controls endothelial differentiation and morphogenesis of cardiac precursors
Erratum in
- Sci Signal. 2012 Oct 9;5(245):er6
-
Erratum for the Research Article "Matrix rigidity controls endothelial differentiation and morphogenesis of cardiac precursors" by Kshitiz et al.Sci Signal. 2023 Aug 29;16(800):eadd8079. doi: 10.1126/scisignal.add8079. Epub 2023 Aug 29. Sci Signal. 2023. PMID: 37643245 No abstract available.
Abstract
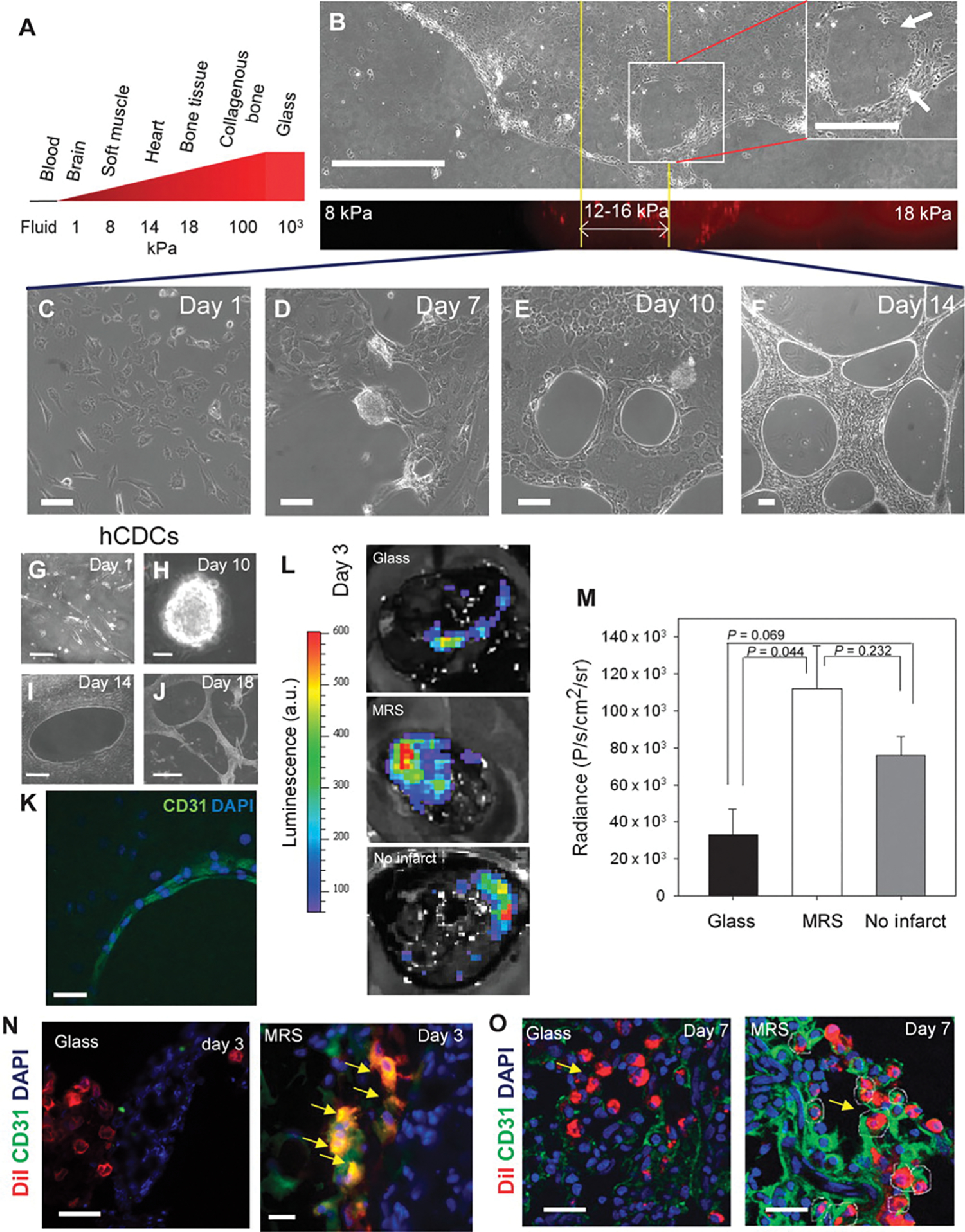
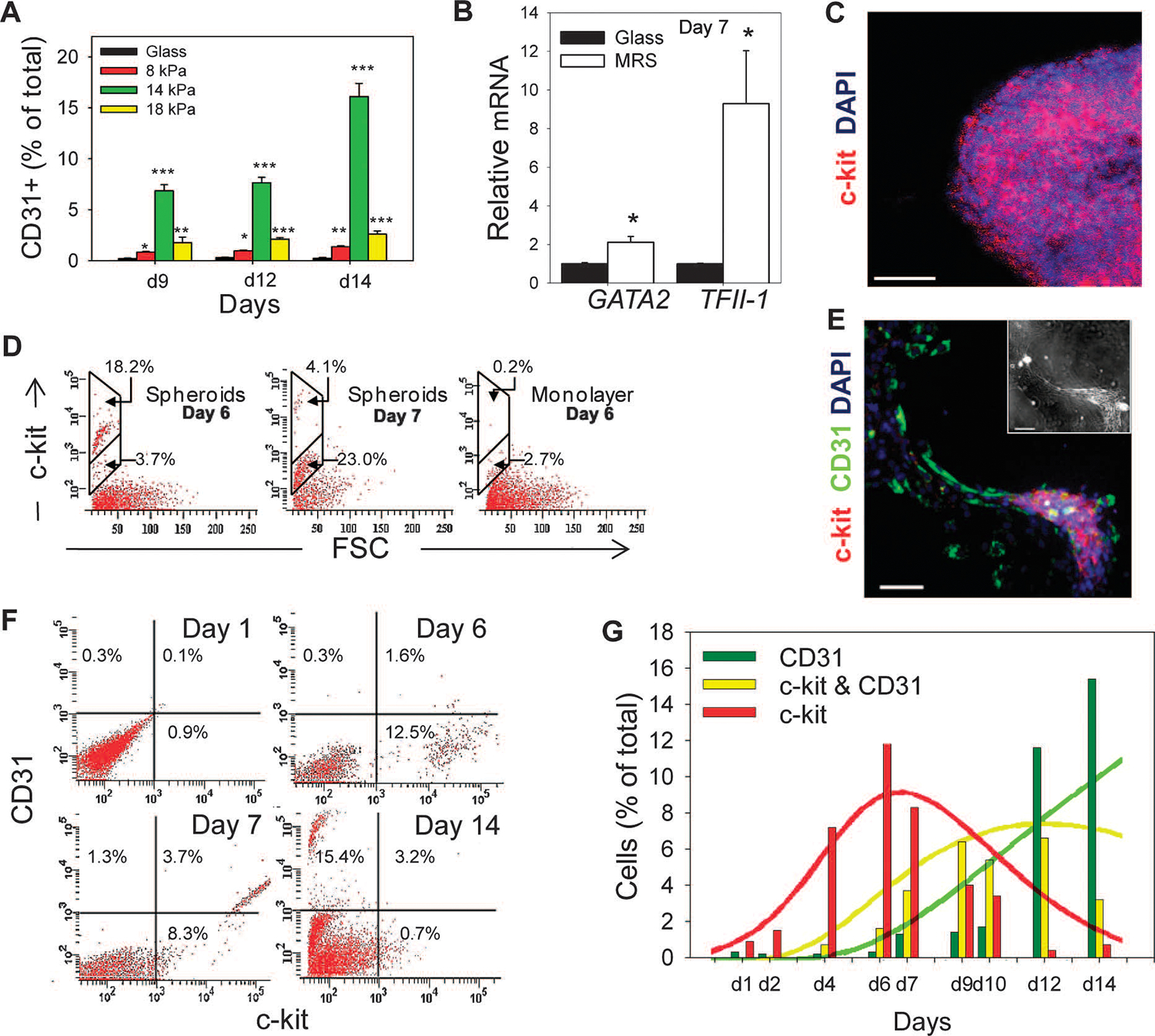
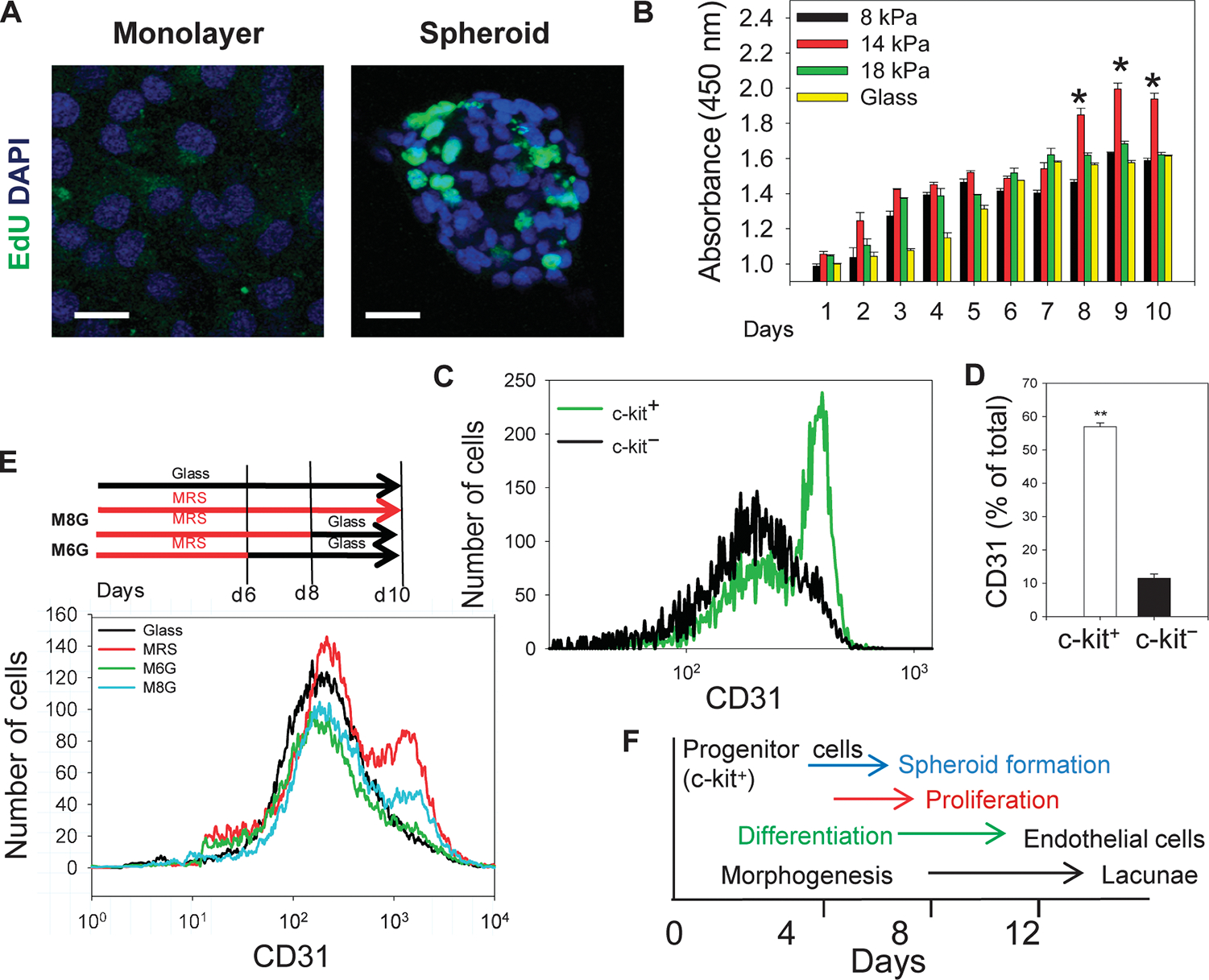
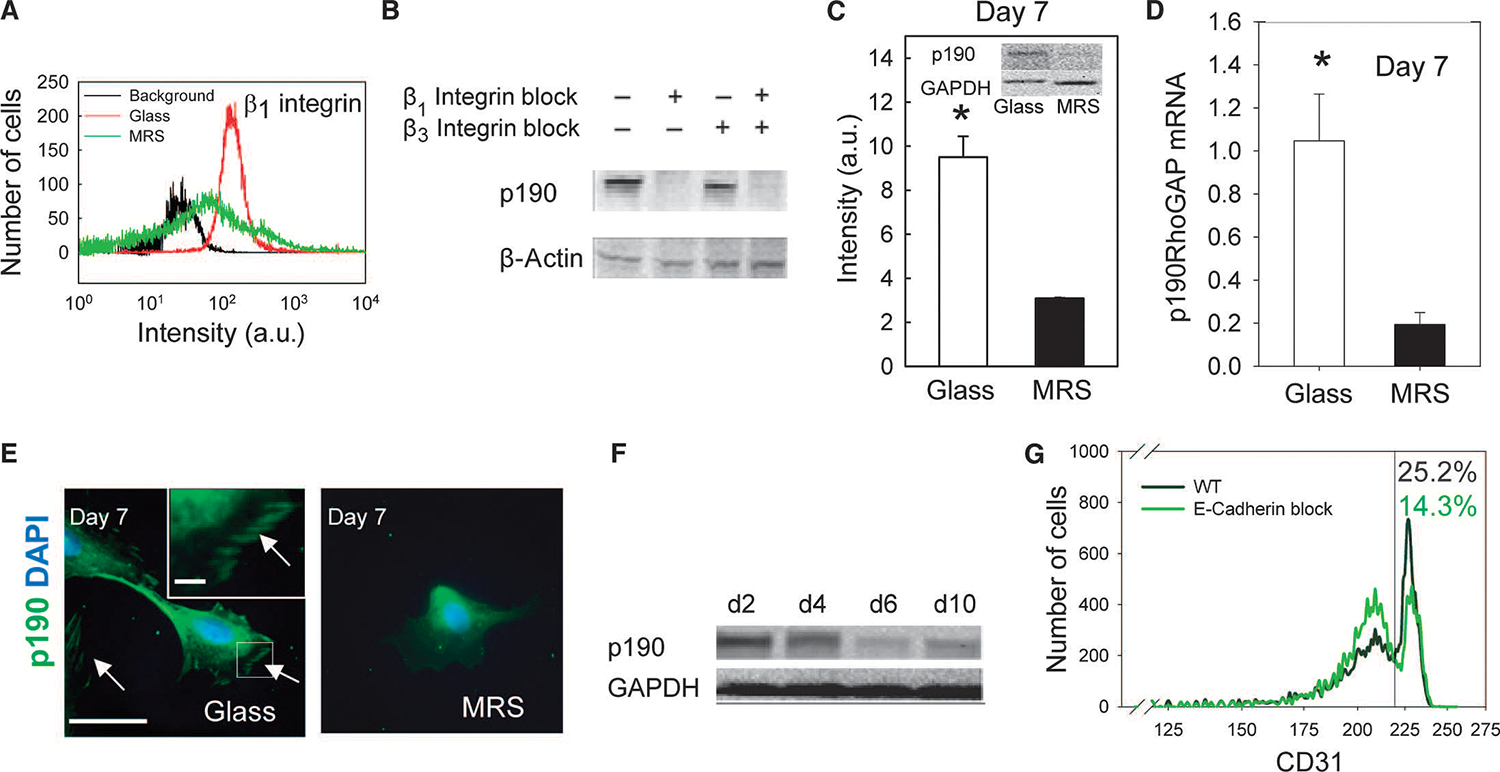
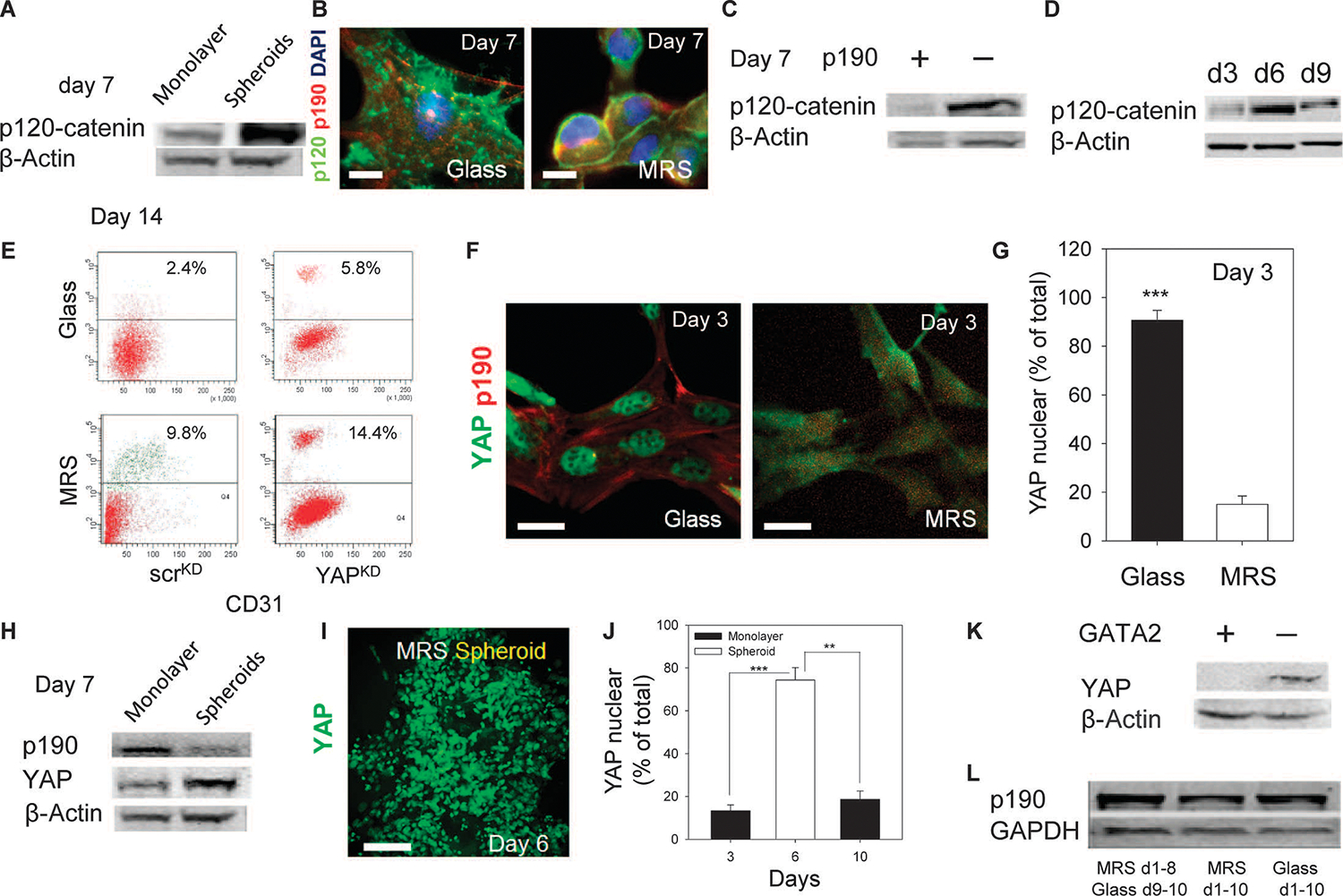
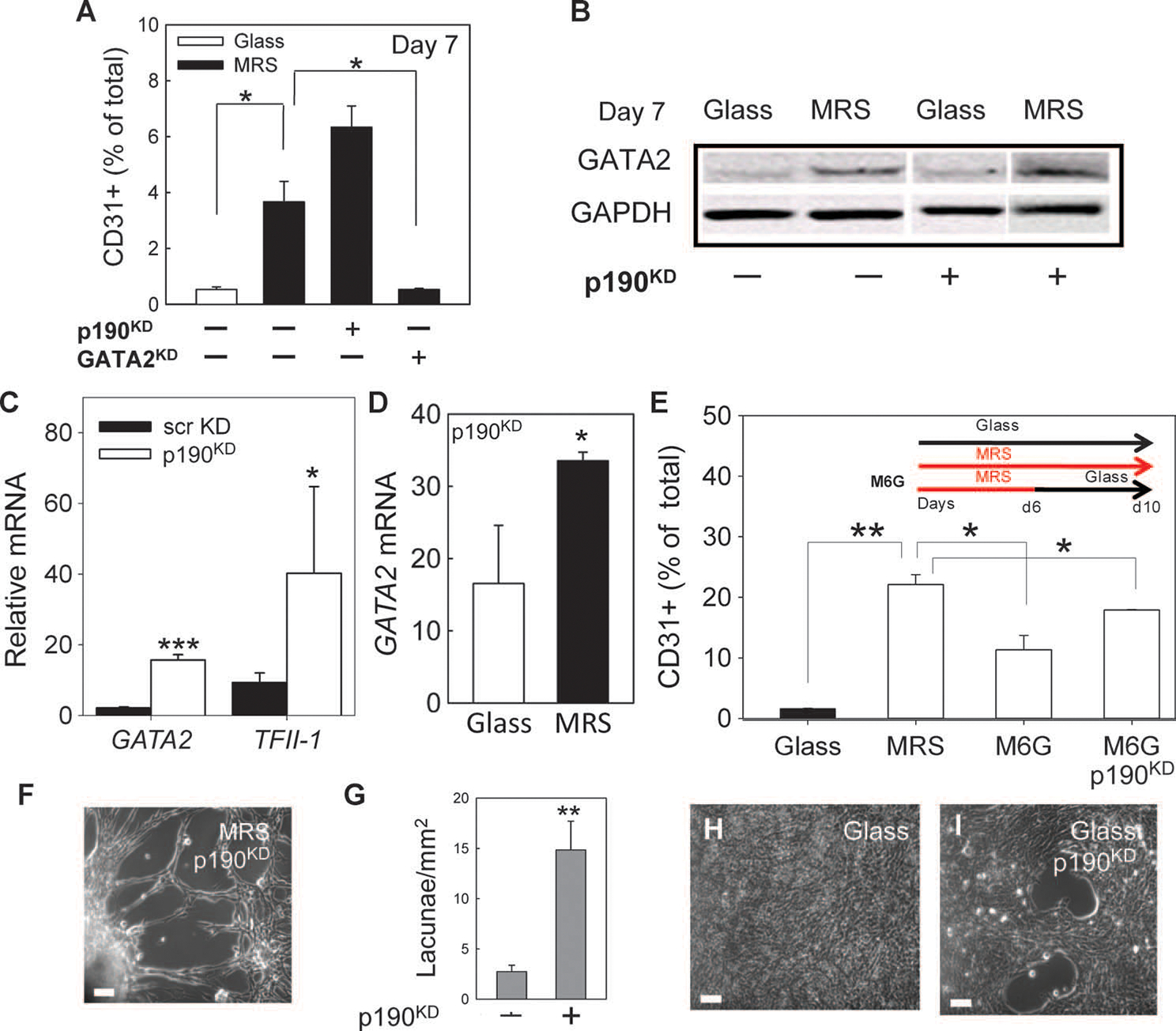
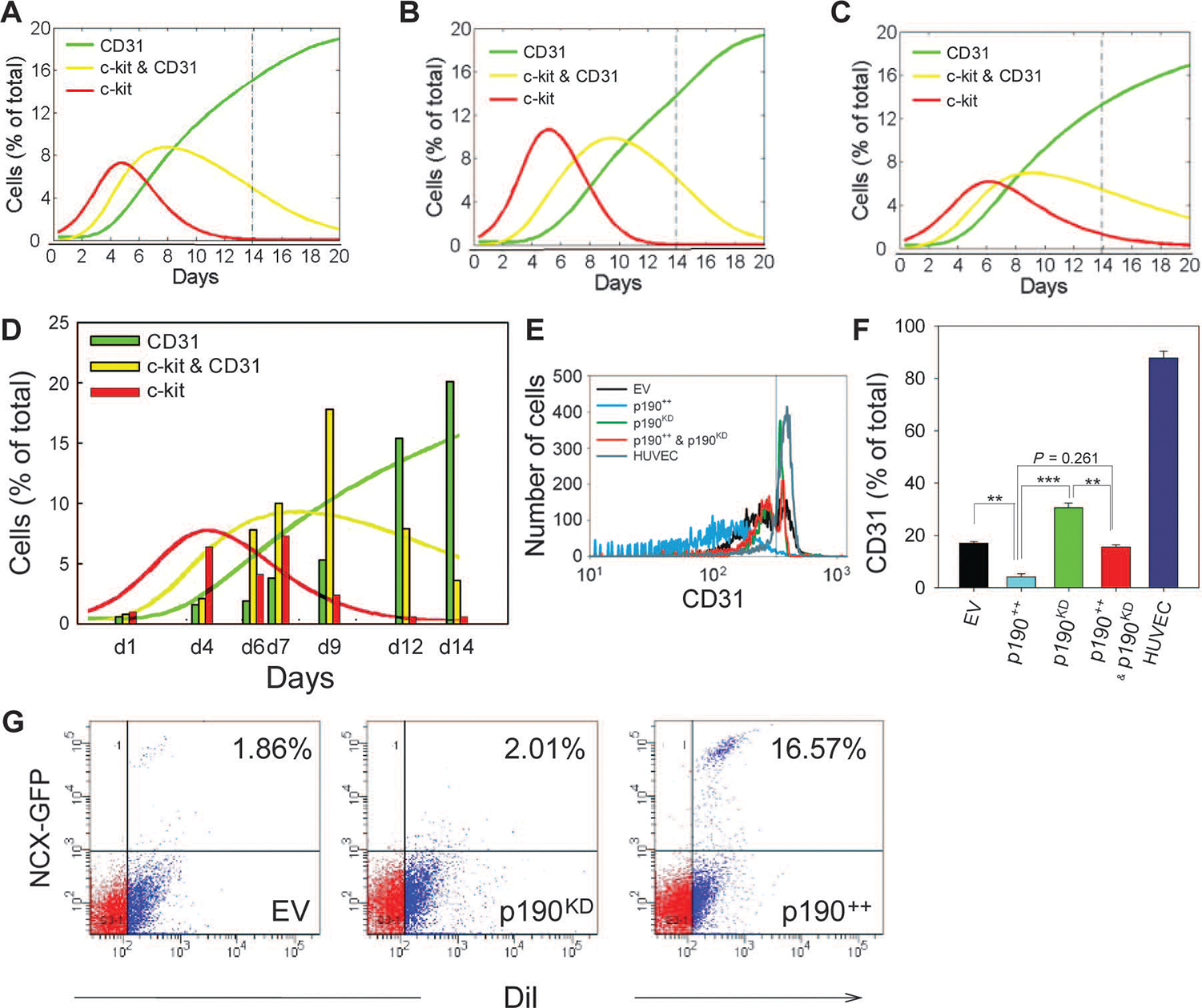
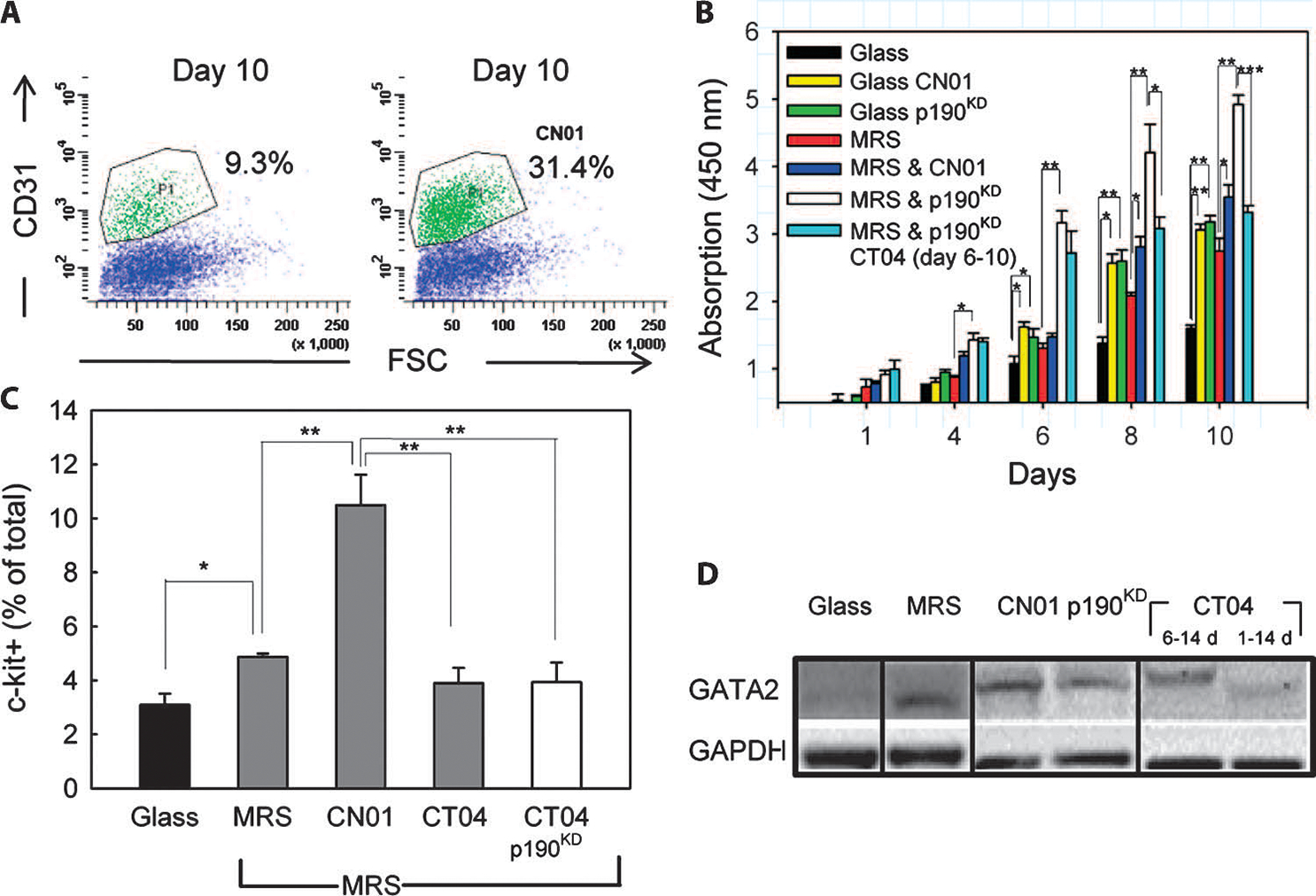
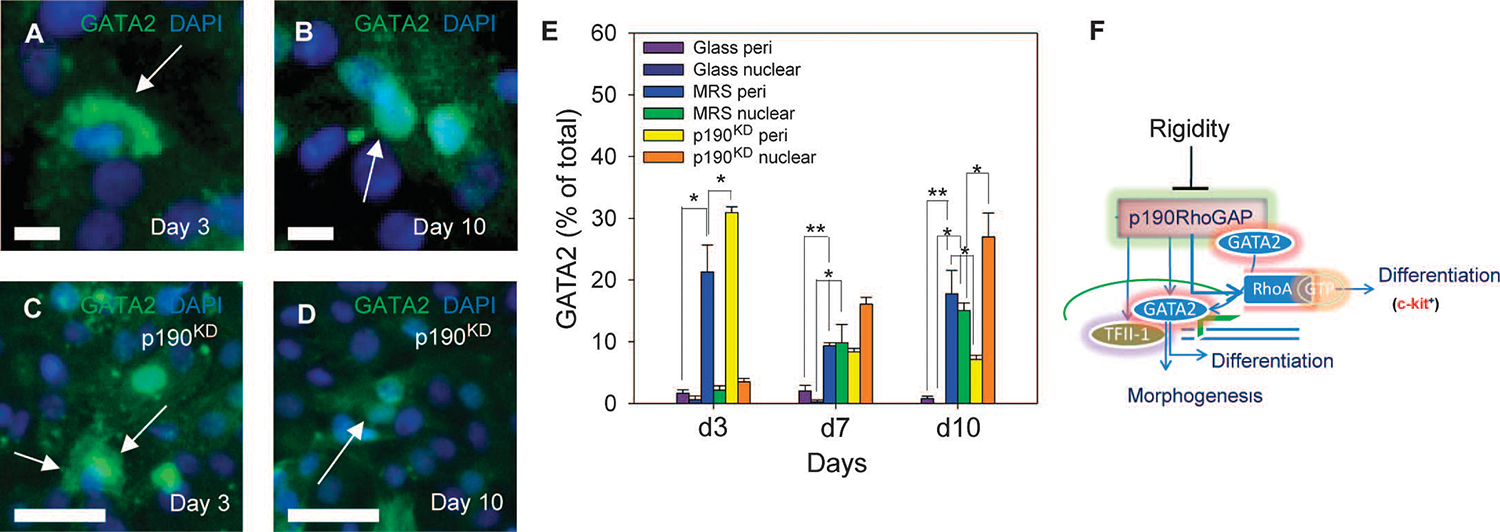
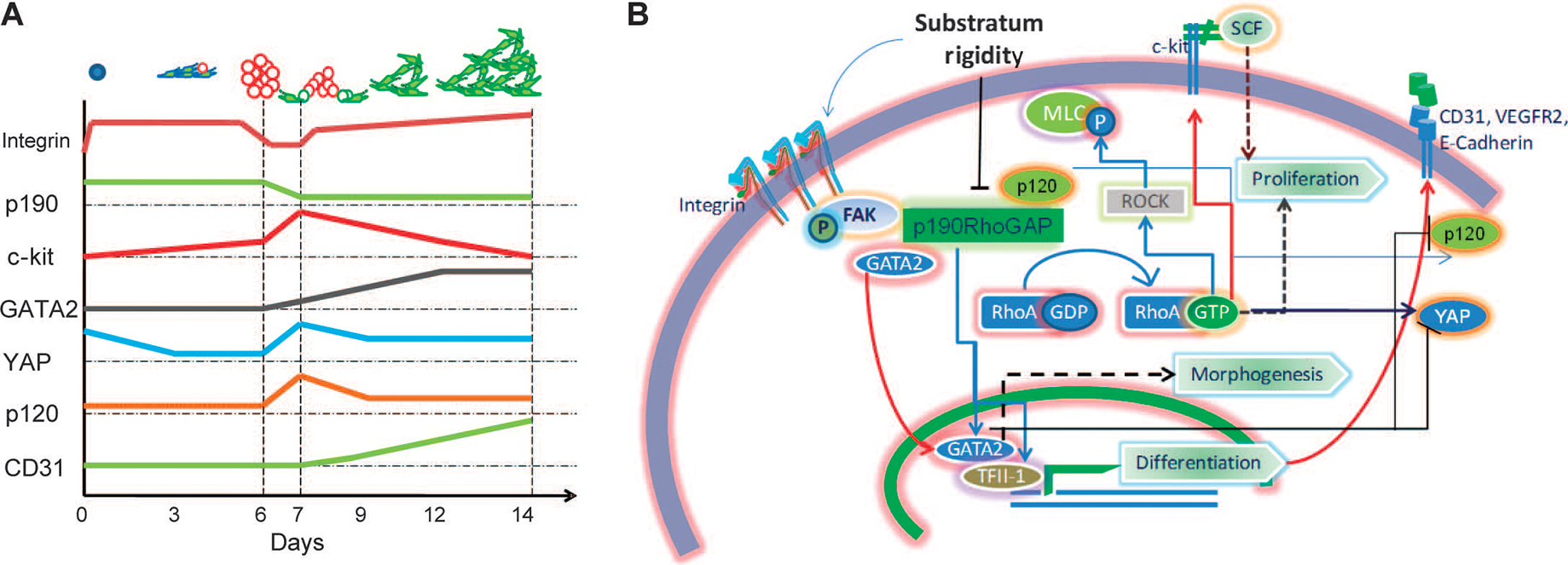
Tissue development and regeneration involve tightly coordinated and integrated processes: selective proliferation of resident stem and precursor cells, differentiation into target somatic cell type, and spatial morphological organization. The role of the mechanical environment in the coordination of these processes is poorly understood. We show that multipotent cells derived from native cardiac tissue continually monitored cell substratum rigidity and showed enhanced proliferation, endothelial differentiation, and morphogenesis when the cell substratum rigidity closely matched that of myocardium. Mechanoregulation of these diverse processes required p190RhoGAP, a guanosine triphosphatase-activating protein for RhoA, acting through RhoA-dependent and -independent mechanisms. Natural or induced decreases in the abundance of p190RhoGAP triggered a series of developmental events by coupling cell-cell and cell-substratum interactions to genetic circuits controlling differentiation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
