

Fine-scale genetic breaks driven by historical range dynamics and ongoing density-barrier effects in the estuarine seaweed Fucus ceranoides L
- PMID: 22672720
- PMCID: PMC3483196
- DOI: 10.1186/1471-2148-12-78
Fine-scale genetic breaks driven by historical range dynamics and ongoing density-barrier effects in the estuarine seaweed Fucus ceranoides L
Abstract
Background: Factors promoting the emergence of sharp phylogeographic breaks include restricted dispersal, habitat discontinuity, physical barriers, disruptive selection, mating incompatibility, genetic surfing and secondary contact. Disentangling the role of each in any particular system can be difficult, especially when species are evenly distributed across transition zones and dispersal barriers are not evident. The estuarine seaweed Fucus ceranoides provides a good example of highly differentiated populations along its most persistent distributional range at the present rear edge of the species distribution, in NW Iberia. Intrinsic dispersal restrictions are obvious in this species, but have not prevented F. ceranoides from vastly expanding its range northwards following the last glaciation, implying that additional factors are responsible for the lack of connectivity between neighbouring southern populations. In this study we analyze 22 consecutive populations of F. ceranoides along NW Iberia to investigate the processes generating and maintaining the observed high levels of regional genetic divergence.
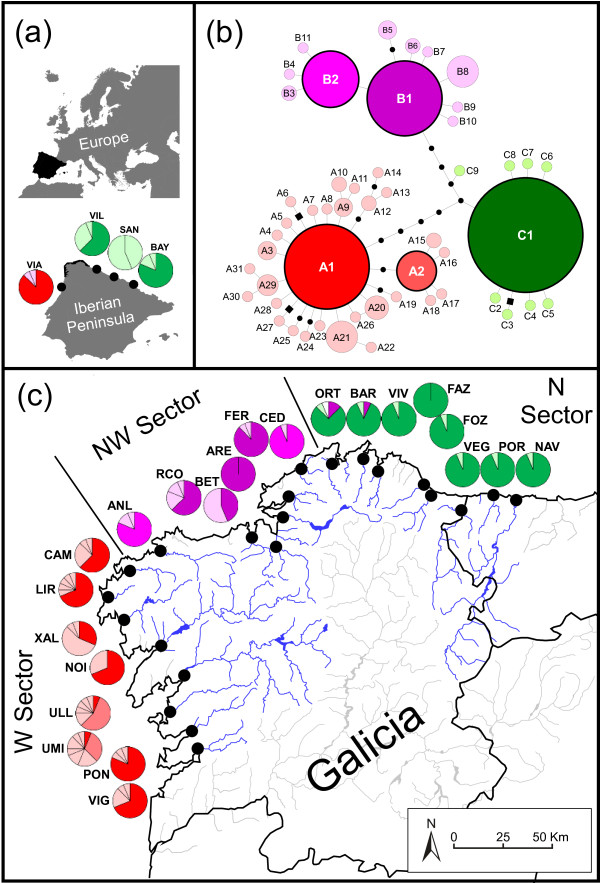
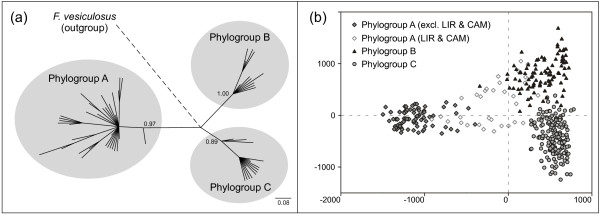
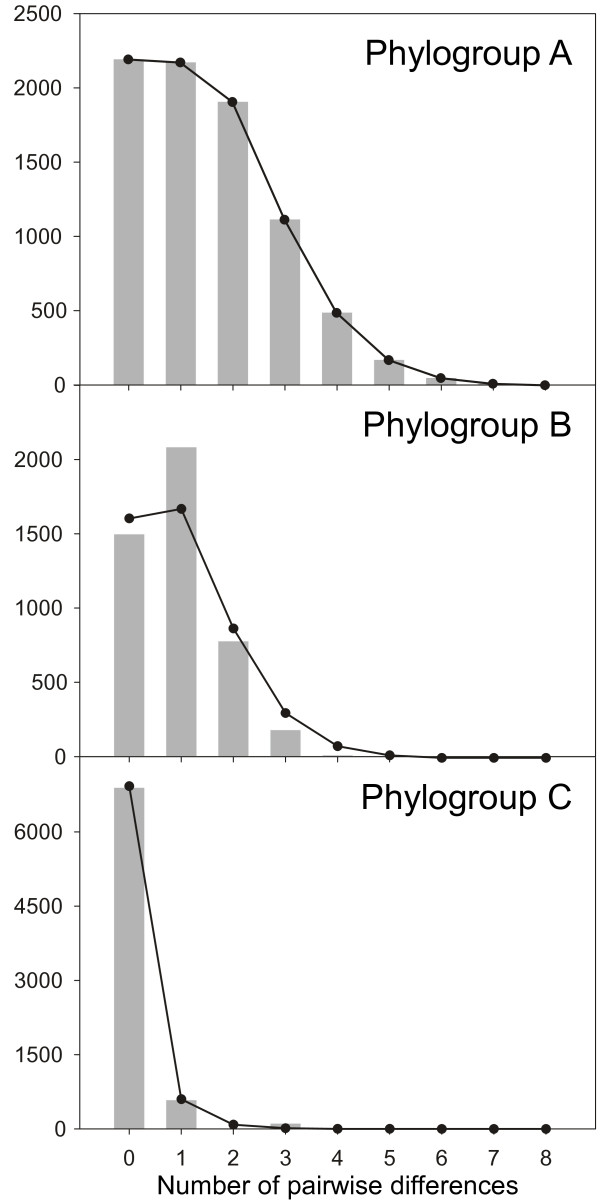
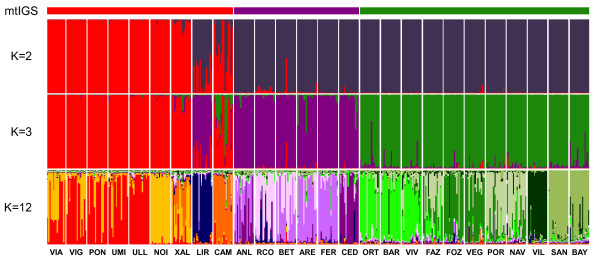
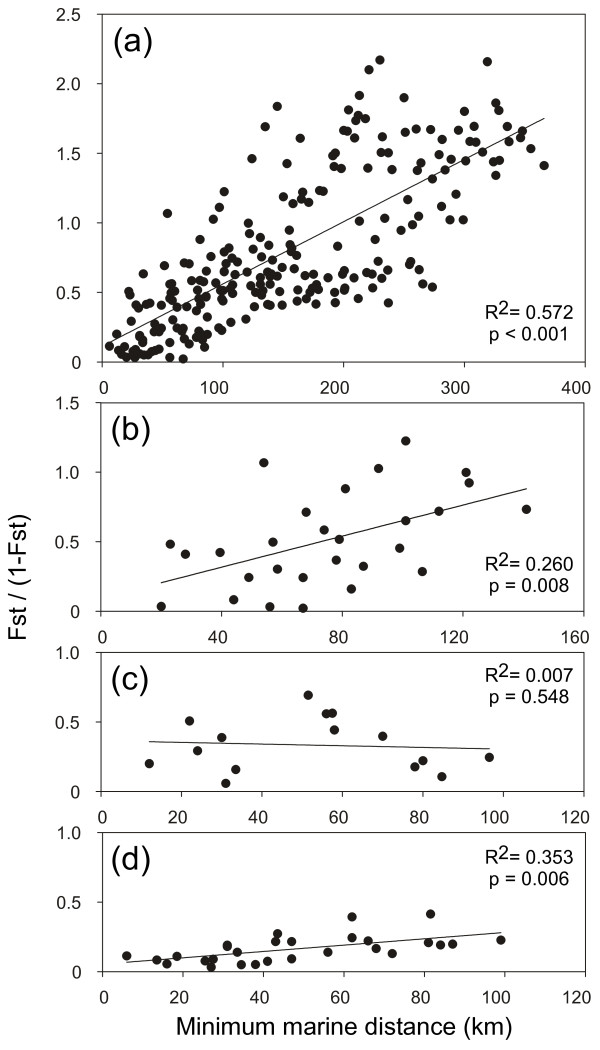
Results: Variation at seven microsatellite loci and at mtDNA spacer sequences was concordant in revealing that Iberian F. ceranoides is composed of three divergent genetic clusters displaying nearly disjunct geographical distributions. Structure and AFC analyses detected two populations with an admixed nuclear background. Haplotypic diversity was high in the W sector and very low in the N sector. Within each genetic cluster, population structure was also pervasive, although shallower.
Conclusions: The deep divergence between sectors coupled with the lack of support for a role of oceanographic barriers in defining the location of breaks suggested 1) that the parapatric genetic sectors result from the regional reassembly of formerly vicariant sub-populations, and 2) that the genetic discontinuities at secondary contact zones (and elsewhere) are maintained despite normal migration rates. We conclude that colonization and immigration, as sources of gene-flow, have very different genetic effects. Migration between established populations is effectively too low to prevent their differentiation by drift or to smooth historical differences inherited from the colonization process. F. ceranoides, but possibly low-dispersal species in general, appear to be unified to a large extent by historical, non-equilibrium processes of extinction and colonization, rather than by contemporary patterns of gene flow.
Figures
Similar articles
-
Surfing the wave on a borrowed board: range expansion and spread of introgressed organellar genomes in the seaweed Fucus ceranoides L.Mol Ecol. 2010 Nov;19(21):4812-22. doi: 10.1111/j.1365-294X.2010.04853.x. Epub 2010 Oct 7. Mol Ecol. 2010. PMID: 20958817
-
Species replacement along a linear coastal habitat: phylogeography and speciation in the red alga Mazzaella laminarioides along the south east Pacific.BMC Evol Biol. 2012 Jun 25;12:97. doi: 10.1186/1471-2148-12-97. BMC Evol Biol. 2012. PMID: 22731925 Free PMC article.
-
Decadal stability in genetic variation and structure in the intertidal seaweed Fucus serratus (Heterokontophyta: Fucaceae).BMC Evol Biol. 2018 Jun 15;18(1):94. doi: 10.1186/s12862-018-1213-2. BMC Evol Biol. 2018. PMID: 29907080 Free PMC article.
-
Population connectivity and phylogeography of a coastal fish, Atractoscion aequidens (Sciaenidae), across the Benguela Current region: evidence of an ancient vicariant event.PLoS One. 2014 Feb 20;9(2):e87907. doi: 10.1371/journal.pone.0087907. eCollection 2014. PLoS One. 2014. PMID: 24586296 Free PMC article.
-
Using genetics to understand the dynamics of wild primate populations.Primates. 2009 Apr;50(2):105-20. doi: 10.1007/s10329-008-0124-z. Epub 2009 Jan 28. Primates. 2009. PMID: 19172380 Free PMC article. Review.
Cited by
-
What maintains the central North Pacific genetic discontinuity in Pacific herring?PLoS One. 2012;7(12):e50340. doi: 10.1371/journal.pone.0050340. Epub 2012 Dec 28. PLoS One. 2012. PMID: 23300525 Free PMC article.
-
Ecological Specialization of Two Photobiont-Specific Maritime Cyanolichen Species of the Genus Lichina.PLoS One. 2015 Jul 16;10(7):e0132718. doi: 10.1371/journal.pone.0132718. eCollection 2015. PLoS One. 2015. PMID: 26181436 Free PMC article.
-
High and distinct range-edge genetic diversity despite local bottlenecks.PLoS One. 2013 Jul 10;8(7):e68646. doi: 10.1371/journal.pone.0068646. eCollection 2013. PLoS One. 2013. PMID: 23967038 Free PMC article.
-
Highly restricted dispersal in habitat-forming seaweed may impede natural recovery of disturbed populations.Sci Rep. 2021 Aug 18;11(1):16792. doi: 10.1038/s41598-021-96027-x. Sci Rep. 2021. PMID: 34408197 Free PMC article.
-
Historical isolation and contemporary gene flow drive population diversity of the brown alga Sargassum thunbergii along the coast of China.BMC Evol Biol. 2017 Dec 7;17(1):246. doi: 10.1186/s12862-017-1089-6. BMC Evol Biol. 2017. PMID: 29216823 Free PMC article.
References
-
- Cheang CC, Chu KH, Ang PO Jr. Phylogeography of the marine macroalgaSargassum hemiphyllum(Phaeophyceae, Heterokontophyta) in northwestern Pacific. Mol Ecol. 2010;19(14):2933–2948. - PubMed
-
- Coyer JA, Hoarau G, Costa JF, Hogerdijk B, Serrao EA, Billard E, Valero M, Pearson GA, Olsen JL. Evolution and diversification within the intertidal brown macroalgaeFucus spiralis/F. vesiculosusspecies complex in the North Atlantic. Mol Phylogenet Evol. 2011;58(2):283–296. - PubMed
-
- Hoarau G, Coyer JA, Veldsink JH, Stam WT, Olsen JL. Glacial refugia and recolonization pathways in the brown seaweedFucus serratus. Mol Ecol. 2007;16(17):3606–3616. - PubMed
-
- Tellier F, Meynard AP, Correa JA, Faugeron S, Valero M. Phylogeographic analyses of the 30 degrees S south-east Pacific biogeographic transition zone establish the occurrence of a sharp genetic discontinuity in the kelpLessonia nigrescens: vicariance or parapatry? Mol Phylogenet Evol. 2009;53(3):679–693. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
