

Potential role of multiple carbon fixation pathways during lipid accumulation in Phaeodactylum tricornutum
- PMID: 22672912
- PMCID: PMC3457861
- DOI: 10.1186/1754-6834-5-40
Potential role of multiple carbon fixation pathways during lipid accumulation in Phaeodactylum tricornutum
Abstract
Background: Phaeodactylum tricornutum is a unicellular diatom in the class Bacillariophyceae. The full genome has been sequenced (<30 Mb), and approximately 20 to 30% triacylglyceride (TAG) accumulation on a dry cell basis has been reported under different growth conditions. To elucidate P. tricornutum gene expression profiles during nutrient-deprivation and lipid-accumulation, cell cultures were grown with a nitrate to phosphate ratio of 20:1 (N:P) and whole-genome transcripts were monitored over time via RNA-sequence determination.
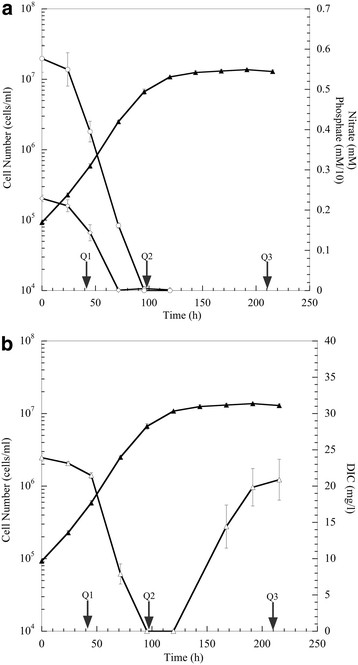
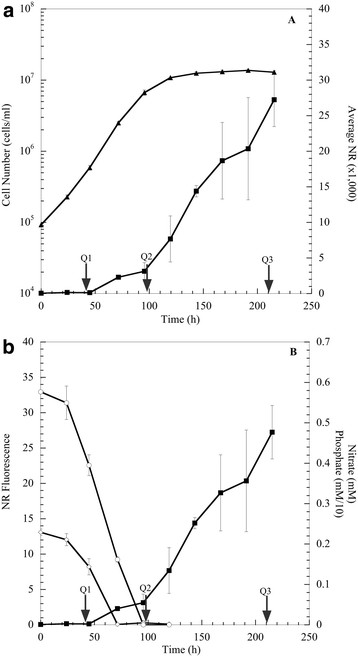
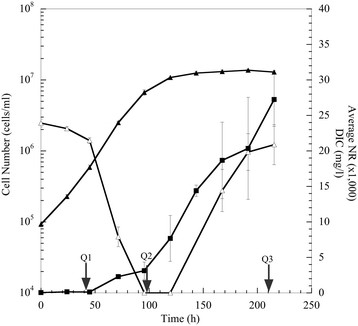
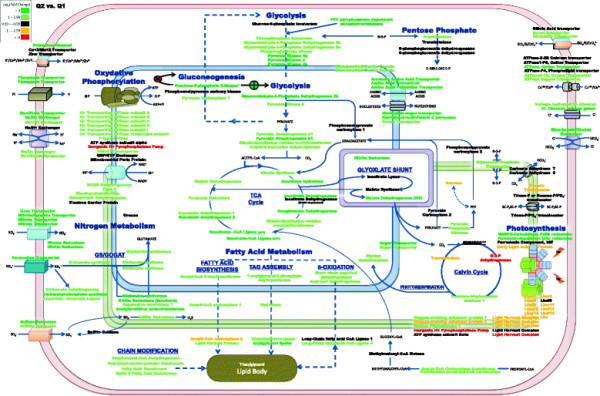
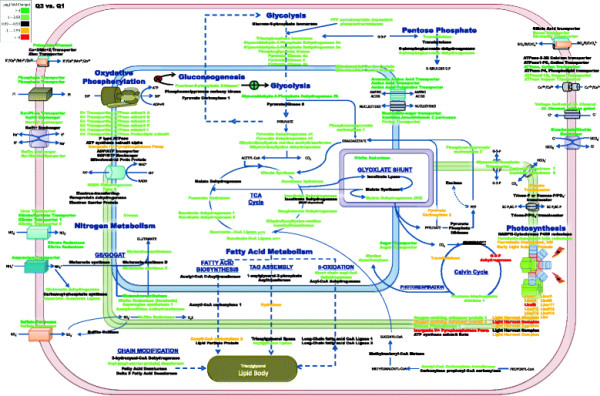
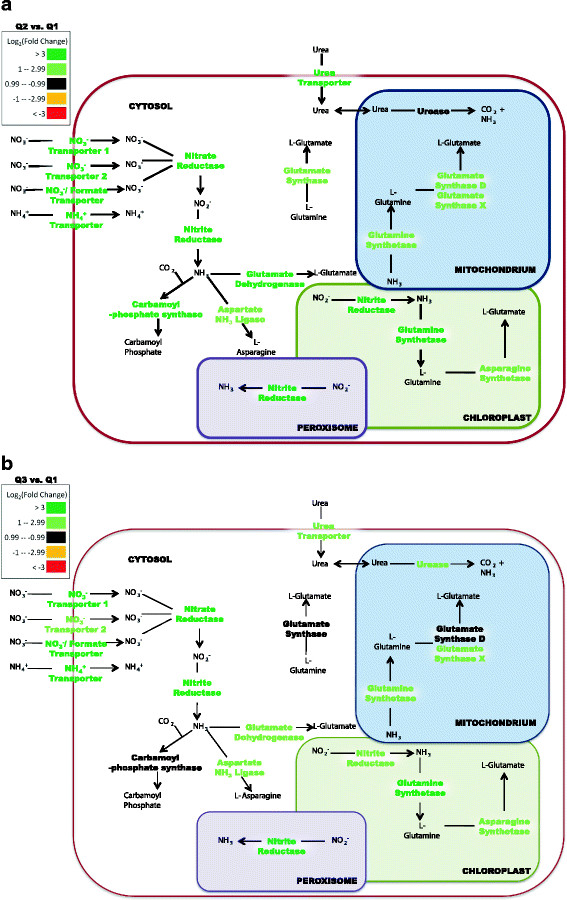
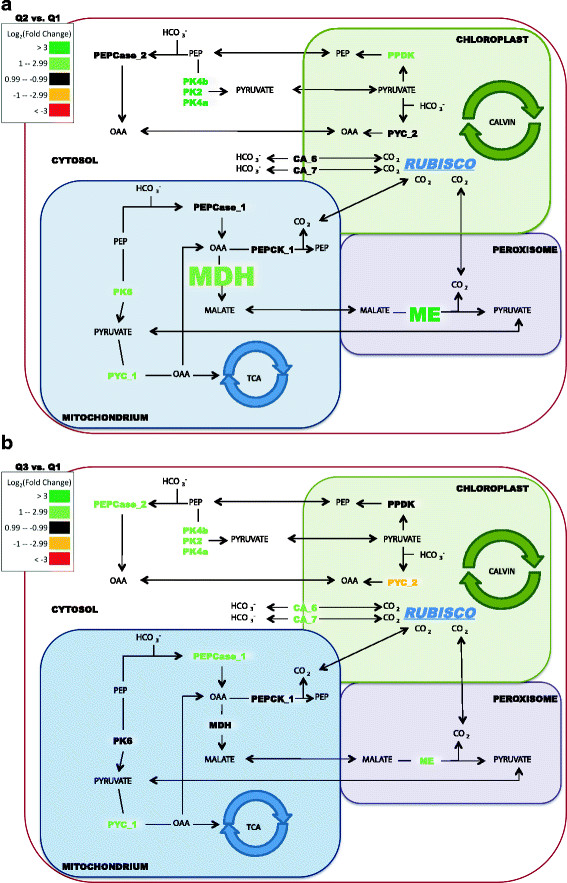
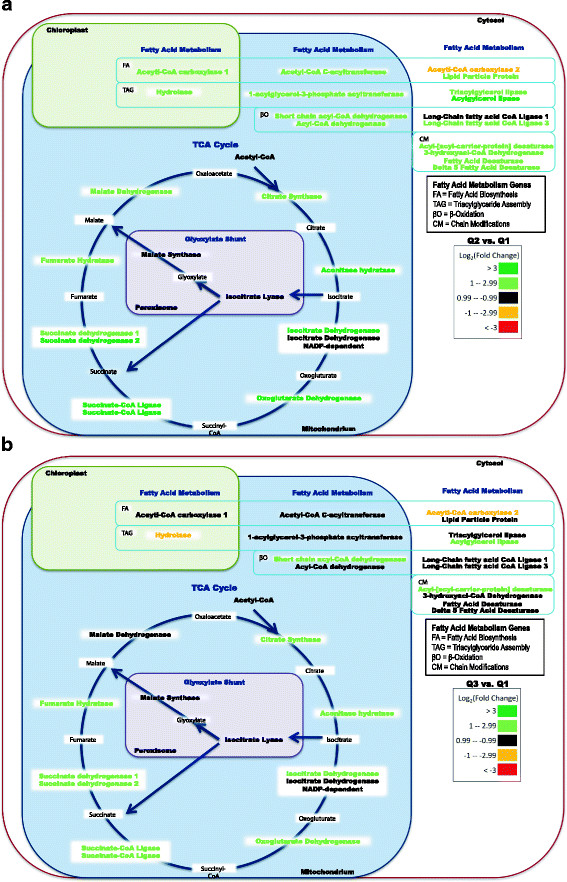
Results: The specific Nile Red (NR) fluorescence (NR fluorescence per cell) increased over time; however, the increase in NR fluorescence was initiated before external nitrate was completely exhausted. Exogenous phosphate was depleted before nitrate, and these results indicated that the depletion of exogenous phosphate might be an early trigger for lipid accumulation that is magnified upon nitrate depletion. As expected, many of the genes associated with nitrate and phosphate utilization were up-expressed. The diatom-specific cyclins cyc7 and cyc10 were down-expressed during the nutrient-deplete state, and cyclin B1 was up-expressed during lipid-accumulation after growth cessation. While many of the genes associated with the C3 pathway for photosynthetic carbon reduction were not significantly altered, genes involved in a putative C4 pathway for photosynthetic carbon assimilation were up-expressed as the cells depleted nitrate, phosphate, and exogenous dissolved inorganic carbon (DIC) levels. P. tricornutum has multiple, putative carbonic anhydrases, but only two were significantly up-expressed (2-fold and 4-fold) at the last time point when exogenous DIC levels had increased after the cessation of growth. Alternative pathways that could utilize HCO3- were also suggested by the gene expression profiles (e.g., putative propionyl-CoA and methylmalonyl-CoA decarboxylases).
Conclusions: The results indicate that P. tricornutum continued carbon dioxide reduction when population growth was arrested and different carbon-concentrating mechanisms were used dependent upon exogenous DIC levels. Based upon overall low gene expression levels for fatty acid synthesis, the results also suggest that the build-up of precursors to the acetyl-CoA carboxylases may play a more significant role in TAG synthesis rather than the actual enzyme levels of acetyl-CoA carboxylases per se. The presented insights into the types and timing of cellular responses to inorganic carbon will help maximize photoautotrophic carbon flow to lipid accumulation.
Figures
Similar articles
-
Nutrient resupplementation arrests bio-oil accumulation in Phaeodactylum tricornutum.Appl Microbiol Biotechnol. 2013 Aug;97(15):7049-59. doi: 10.1007/s00253-013-5010-y. Epub 2013 Jun 15. Appl Microbiol Biotechnol. 2013. PMID: 23771779 Free PMC article.
-
Effects and mechanisms of glyphosate as phosphorus nutrient on element stoichiometry and metabolism in the diatom Phaeodactylum tricornutum.Appl Environ Microbiol. 2024 Feb 21;90(2):e0213123. doi: 10.1128/aem.02131-23. Epub 2024 Jan 24. Appl Environ Microbiol. 2024. PMID: 38265214 Free PMC article.
-
Proteomic and biochemical responses to different concentrations of CO2 suggest the existence of multiple carbon metabolism strategies in Phaeodactylum tricornutum.Biotechnol Biofuels. 2021 Dec 14;14(1):235. doi: 10.1186/s13068-021-02088-5. Biotechnol Biofuels. 2021. PMID: 34906223 Free PMC article.
-
Phaeodactylum tricornutum microalgae as a rich source of omega-3 oil: Progress in lipid induction techniques towards industry adoption.Food Chem. 2019 Nov 1;297:124937. doi: 10.1016/j.foodchem.2019.06.004. Epub 2019 Jun 3. Food Chem. 2019. PMID: 31253257 Review.
-
Amino Acid Catabolism During Nitrogen Limitation in Phaeodactylum tricornutum.Front Plant Sci. 2020 Dec 17;11:589026. doi: 10.3389/fpls.2020.589026. eCollection 2020. Front Plant Sci. 2020. PMID: 33408729 Free PMC article. Review.
Cited by
-
Gene expression studies for the analysis of domoic acid production in the marine diatom Pseudo-nitzschia multiseries.BMC Mol Biol. 2013 Nov 1;14:25. doi: 10.1186/1471-2199-14-25. BMC Mol Biol. 2013. PMID: 24180290 Free PMC article.
-
A rare Phaeodactylum tricornutum cruciform morphotype: culture conditions, transformation and unique fatty acid characteristics.PLoS One. 2014 Apr 7;9(4):e93922. doi: 10.1371/journal.pone.0093922. eCollection 2014. PLoS One. 2014. PMID: 24710200 Free PMC article.
-
Biochemical Characterization of a Novel Redox-Regulated Metacaspase in a Marine Diatom.Front Microbiol. 2021 Sep 8;12:688199. doi: 10.3389/fmicb.2021.688199. eCollection 2021. Front Microbiol. 2021. PMID: 34566902 Free PMC article.
-
Multi-omic analysis on the molecular mechanisms of rapid growth in 'Deqin' alfalfa after space mutagenesis.BMC Plant Biol. 2025 Jan 9;25(1):34. doi: 10.1186/s12870-025-06060-5. BMC Plant Biol. 2025. PMID: 39780091 Free PMC article.
-
Agrigenomics for microalgal biofuel production: an overview of various bioinformatics resources and recent studies to link OMICS to bioenergy and bioeconomy.OMICS. 2013 Nov;17(11):537-49. doi: 10.1089/omi.2013.0025. Epub 2013 Sep 17. OMICS. 2013. PMID: 24044362 Free PMC article. Review.
References
-
- Brown LR. Plan B 2.0: Rescuing a Planet Under Stress and a Civilization in Trouble. Updated and Expanded. WW Norton and Company. 2006.
-
- Dukes JS. Burning buried sunshine: Human consumption of ancient solar energy. Climatic Change. 2003;61:31–44. doi: 10.1023/A:1026391317686. - DOI
-
- Field CB. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science. 1998;281:237–240. - PubMed
-
- Granum E, Raven JA, Leegood RC. How do marine diatoms fix 10 billion tonnes of inorganic carbon per year? Can. 2005;83:898–908. doi: 10.1139/b05-077. - DOI
-
- Falkowski PG, Raven JA. Aquatic. Photosynthesis. 1997.
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
