

A-current and type I/type II transition determine collective spiking from common input
- PMID: 22673330
- PMCID: PMC3544951
- DOI: 10.1152/jn.00928.2011
A-current and type I/type II transition determine collective spiking from common input
Abstract
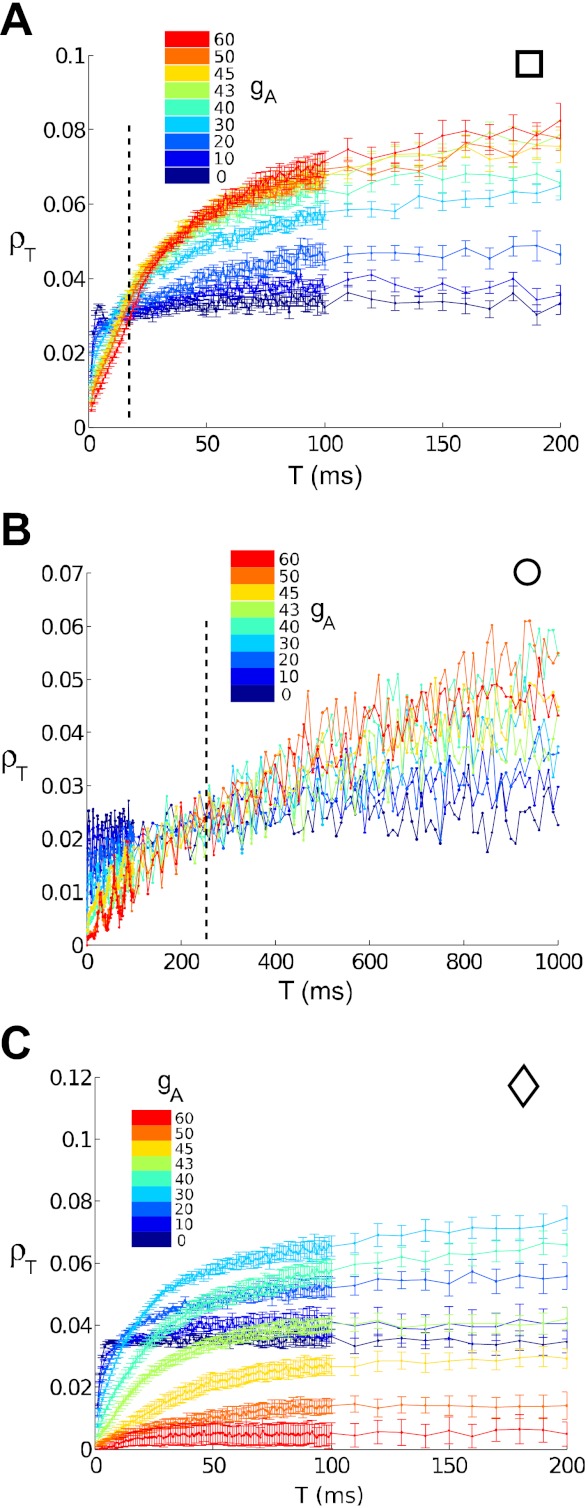
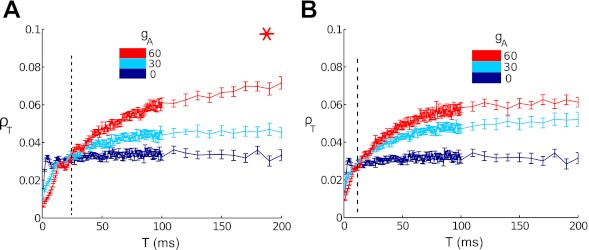
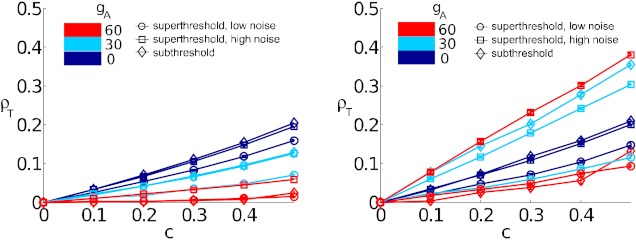
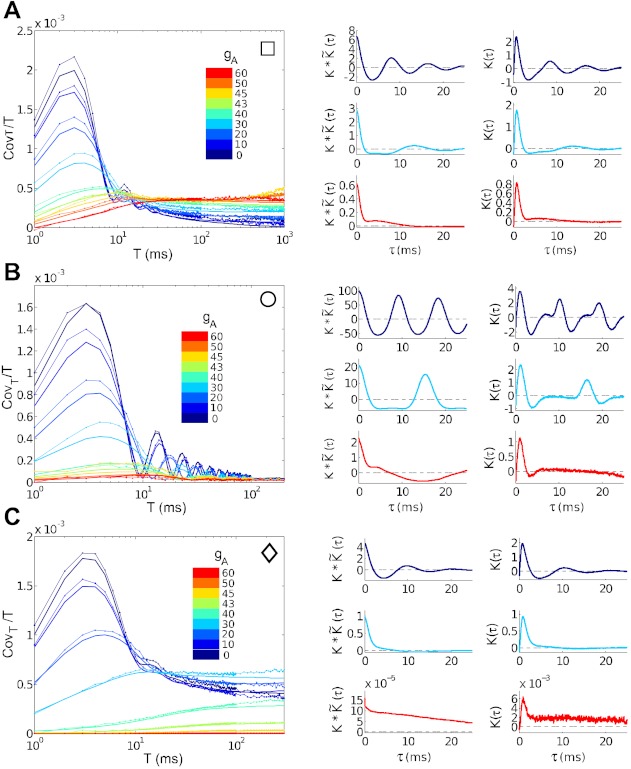
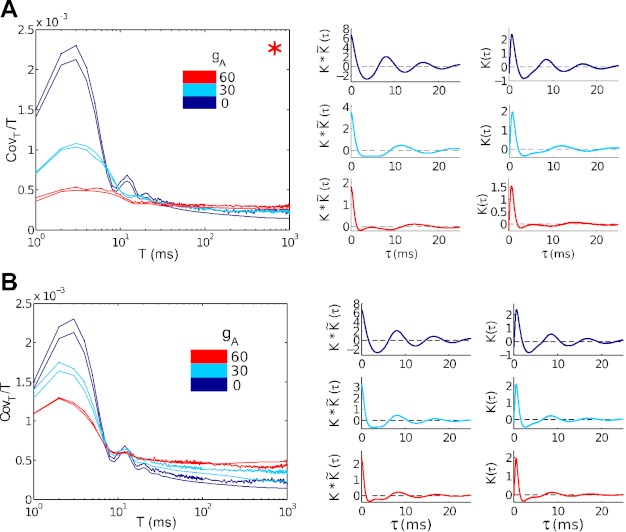
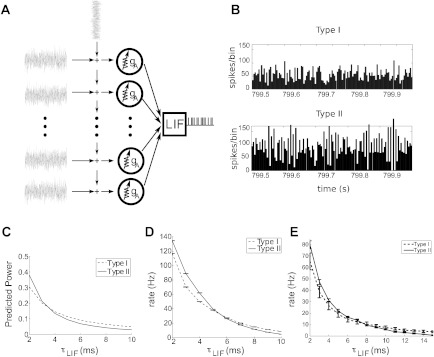
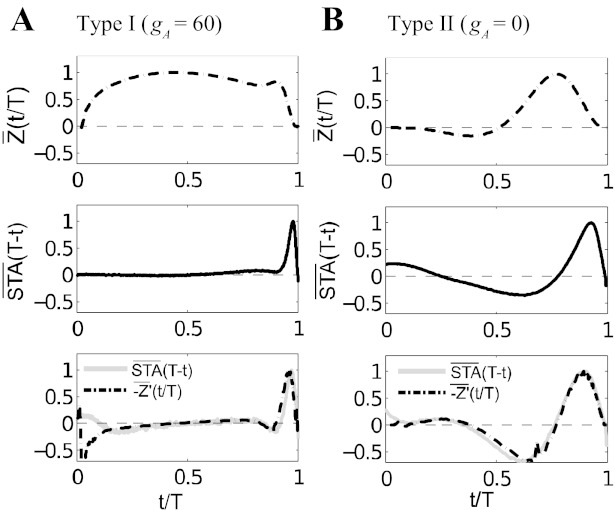
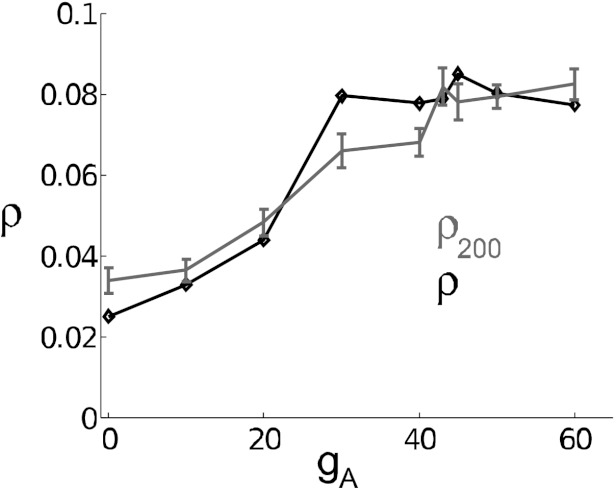
The mechanisms and impact of correlated, or synchronous, firing among pairs and groups of neurons are under intense investigation throughout the nervous system. A ubiquitous circuit feature that can give rise to such correlations consists of overlapping, or common, inputs to pairs and populations of cells, leading to common spike train responses. Here, we use computational tools to study how the transfer of common input currents into common spike outputs is modulated by the physiology of the recipient cells. We focus on a key conductance, g(A), for the A-type potassium current, which drives neurons between "type II" excitability (low g(A)), and "type I" excitability (high g(A)). Regardless of g(A), cells transform common input fluctuations into a tendency to spike nearly simultaneously. However, this process is more pronounced at low g(A) values. Thus, for a given level of common input, type II neurons produce spikes that are relatively more correlated over short time scales. Over long time scales, the trend reverses, with type II neurons producing relatively less correlated spike trains. This is because these cells' increased tendency for simultaneous spiking is balanced by an anticorrelation of spikes at larger time lags. These findings extend and interpret prior findings for phase oscillators to conductance-based neuron models that cover both oscillatory (superthreshold) and subthreshold firing regimes. We demonstrate a novel implication for neural signal processing: downstream cells with long time constants are selectively driven by type I cell populations upstream and those with short time constants by type II cell populations. Our results are established via high-throughput numerical simulations and explained via the cells' filtering properties and nonlinear dynamics.
Figures
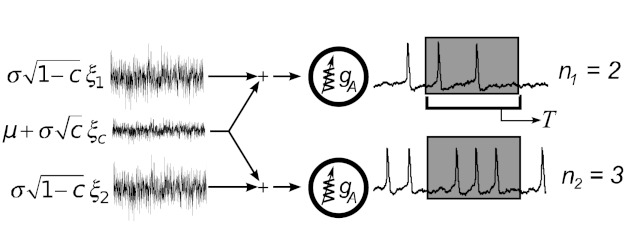
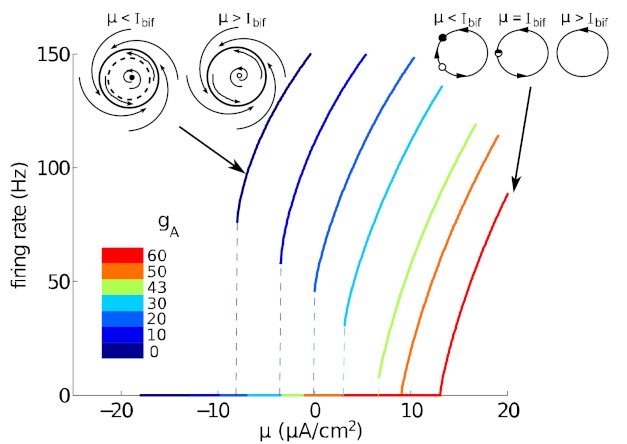
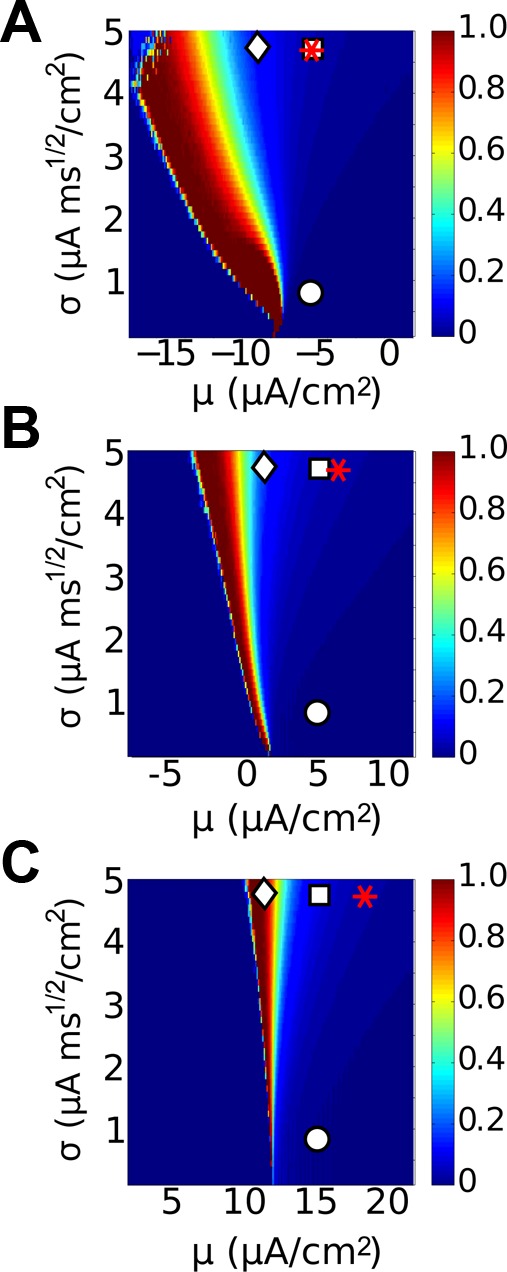
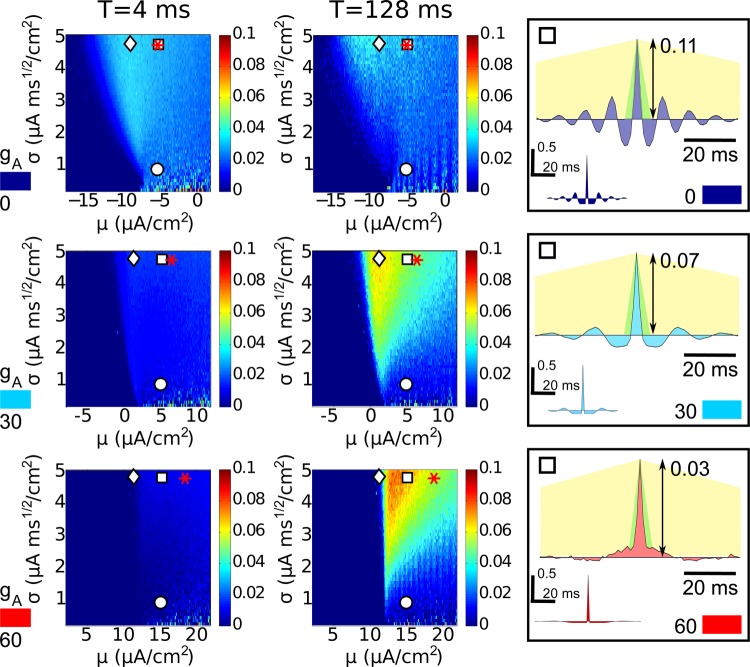
References
-
- Abouzeid A, Ermentrout B. Correlation transfer in stochastically driven neural oscillators over short and long time scales. Phys Rev E Stat Nonlin Soft Matter Phys 84: 061914 - PubMed
-
- Agüera y Arcas B, Fairhall A, Bialek W. Computation in a single neuron: Hodgkin- Huxley revisted. Neural Comput 15: 1715–1749, 2003 - PubMed
-
- Alonso J, Usrey W, Reid W. Precisely correlated firing in cells of the lateral geniculate nucleus Nature 383: 815–819, 1996 - PubMed
-
- Averbeck BB, Latham PE, Pouget A. Neural correlations, population coding, computation. Nat Rev Neuroscience 7: 358–366, 2006 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources