

Prolactin regulates tuberoinfundibular dopamine neuron discharge pattern: novel feedback control mechanisms in the lactotrophic axis
- PMID: 22674282
- PMCID: PMC6620951
- DOI: 10.1523/JNEUROSCI.0129-12.2012
Prolactin regulates tuberoinfundibular dopamine neuron discharge pattern: novel feedback control mechanisms in the lactotrophic axis
Abstract
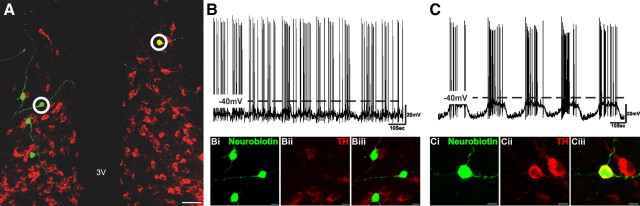
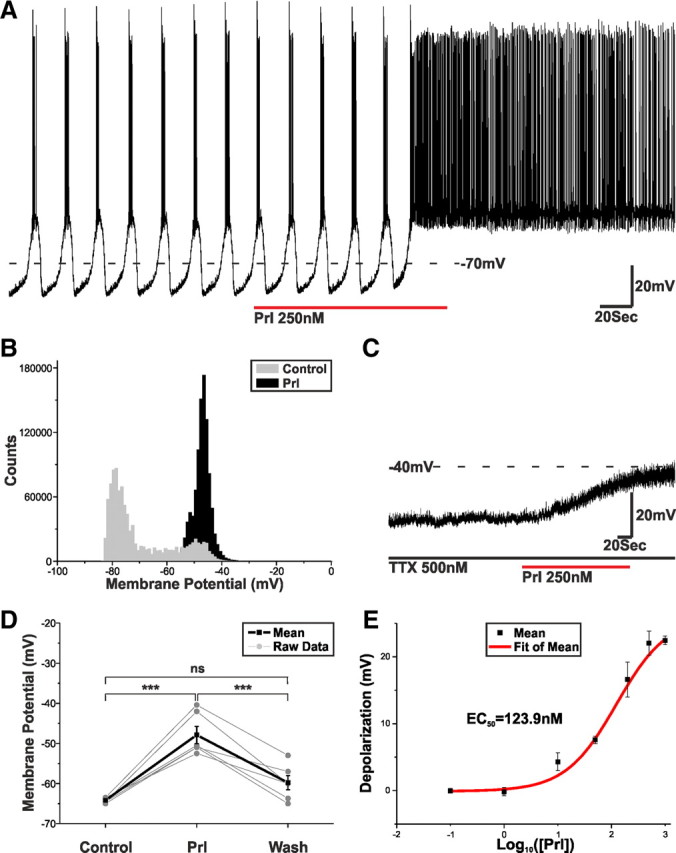
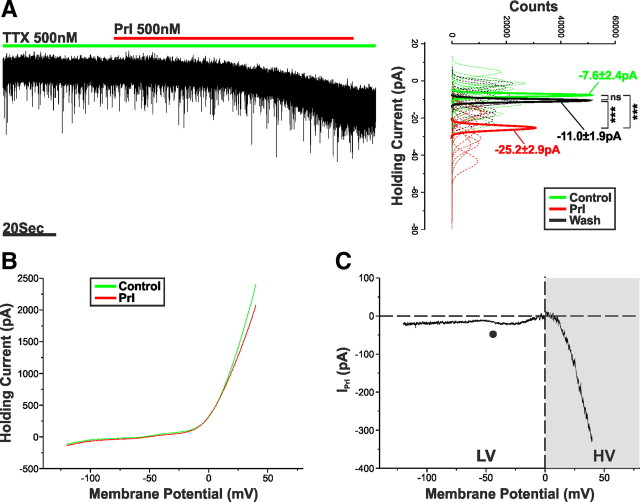
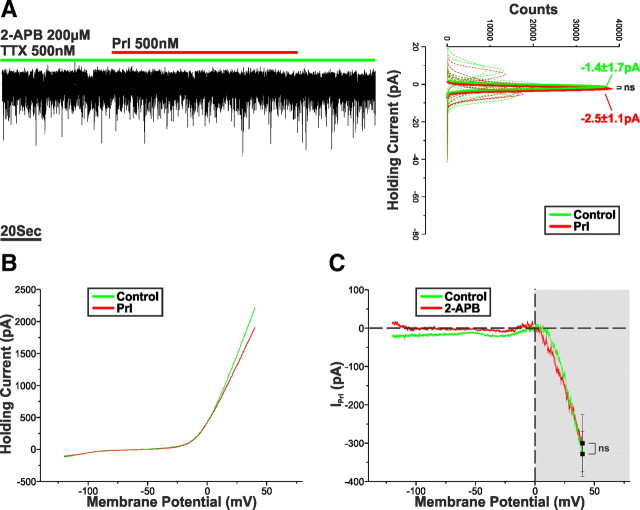
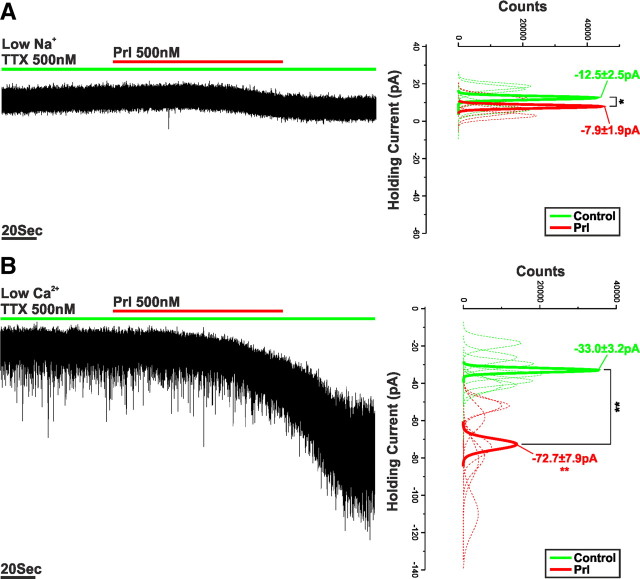
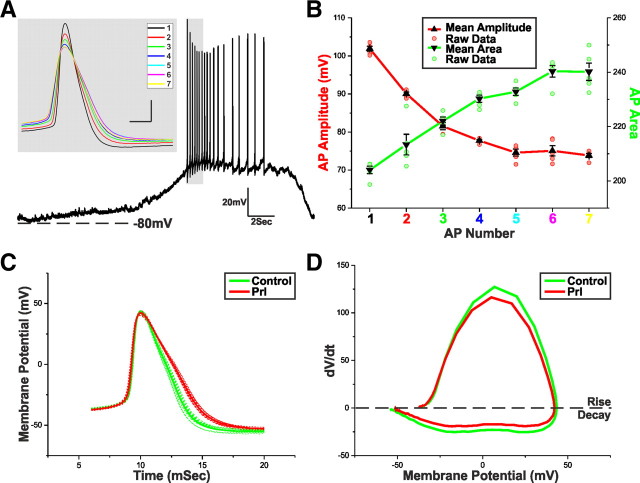
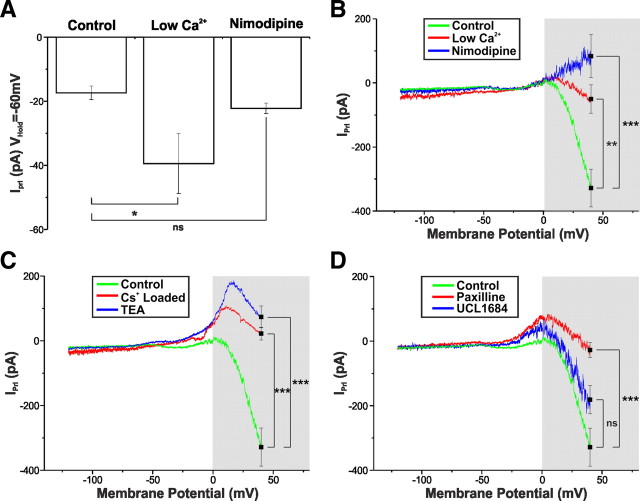
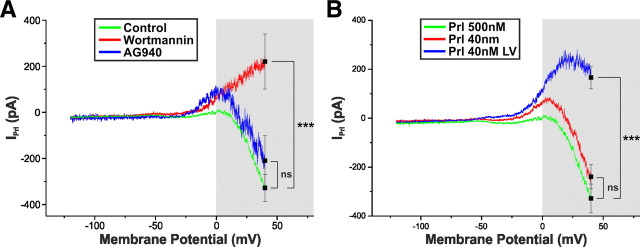
Balance in the body's hormonal axes depends on feedback onto neuroendocrine hypothalamic neurons. This phenomenon involves transcriptional and biosynthetic effects, yet less is known about the potential rapid modulation of electrical properties. Here, we investigated this issue in the lactotrophic axis, in which the pituitary hormone prolactin is tonically inhibited by tuberoinfundibular dopamine (TIDA) neurons located in the hypothalamic arcuate nucleus. Whole-cell recordings were performed on slices of the rat hypothalamus. In the presence of prolactin, spontaneously oscillating TIDA cells depolarized, switched from phasic to tonic discharge, and exhibited broadened action potentials. The underlying prolactin-induced current is composed of separate low- and high-voltage components that include the activation of a transient receptor potential-like current and the inhibition of a Ca(2+)-dependent BK-type K(+) current, respectively, as revealed by ion substitution experiments and pharmacological manipulation. The two components of the prolactin-induced current appear to be mediated through distinct signaling pathways as the high-voltage component is abolished by the phosphoinositide 3-kinase blocker wortmannin, whereas the low-voltage component is not. This first description of the central electrophysiological actions of prolactin suggests a novel feedback mechanism. By simultaneously enhancing the discharge and spike duration of TIDA cells, increased serum prolactin can promote dopamine release to limit its own secretion with implications for the control of lactation, sexual libido, fertility, and body weight.
Figures
References
-
- al-Sakkaf KA, Dobson PR, Brown BL. Activation of phosphatidylinositol 3-kinase by prolactin in Nb2 cells. Biochem Biophys Res Commun. 1996;221:779–784. - PubMed
-
- Amenomori Y, Chen CL, Meites J. Serum prolactin levels in rats during different reproductive states. Endocrinology. 1970;86:506–510. - PubMed
-
- Andrews ZB, Kokay IC, Grattan DR. Dissociation of prolactin secretion from tuberoinfundibular dopamine activity in late pregnant rats. Endocrinology. 2001;142:2719–2724. - PubMed
-
- Arbogast LA, Voogt JL. Hyperprolactinemia increases and hypoprolactinemia decreases tyrosine hydroxylase messenger ribonucleic acid levels in the arcuate nuclei, but not the substantia nigra or zona incerta. Endocrinology. 1991;128:997–1005. - PubMed
-
- Arbogast LA, Voogt JL. Hypoprolactinemia decreases tyrosine hydroxylase activity in the tuberoinfundibular dopaminergic neurons acutely by protein dephosphorylation and chronically by changes in gene expression. Endocrine. 1995;3:801–806. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous