

Experience-dependent expression of NPAS4 regulates plasticity in adult visual cortex
- PMID: 22674715
- PMCID: PMC3487036
- DOI: 10.1113/jphysiol.2012.234237
Experience-dependent expression of NPAS4 regulates plasticity in adult visual cortex
Abstract
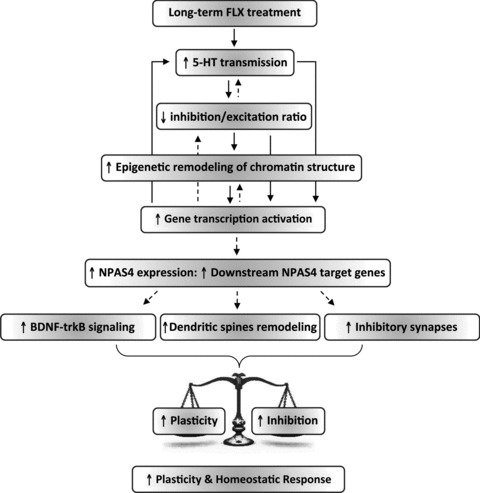
There is evidence that developmental-like plasticity can be reactivated in the adult visual cortex. Although activity-dependent transcription factors underlying the process of plasticity reactivation are currently unknown, recent studies point towards NPAS4 as a candidate gene for the occurrence of plasticity in the adult visual system. Here, we addressed whether NPAS4 is involved in the reinstatement of plasticity by using the monocular deprivation protocol and long-term fluoxetine treatment as a pharmacological strategy that restores plasticity in adulthood. A combination of molecular assays for gene expression and epigenetic analysis, gene delivery by lentiviral infection, shRNA interference and electrophysiology as a functional read-out, revealed a previously unknown role for the transcription factor NPAS4 in the regulation of adult visual cortical plasticity. We found that NPAS4 overexpression restores ocular dominance plasticity in adult naıve animals whereas NPAS4 down-regulation prevents the plastic outcome caused by fluoxetine in adulthood.Our findings lead the way to the identification of novel therapeutic targets for pathological conditions where reorganization of neuronal networks would be beneficial in adult life.
Figures




Similar articles
-
Serotonin triggers a transient epigenetic mechanism that reinstates adult visual cortex plasticity in rats.Eur J Neurosci. 2011 Jan;33(1):49-57. doi: 10.1111/j.1460-9568.2010.07488.x. Epub 2010 Dec 12. Eur J Neurosci. 2011. PMID: 21156002
-
The antidepressant fluoxetine restores plasticity in the adult visual cortex.Science. 2008 Apr 18;320(5874):385-8. doi: 10.1126/science.1150516. Science. 2008. PMID: 18420937
-
Gene expression patterns underlying the reinstatement of plasticity in the adult visual system.Neural Plast. 2013;2013:605079. doi: 10.1155/2013/605079. Epub 2013 Jun 26. Neural Plast. 2013. PMID: 23936678 Free PMC article.
-
Activity-dependent NPAS4 expression and the regulation of gene programs underlying plasticity in the central nervous system.Neural Plast. 2013;2013:683909. doi: 10.1155/2013/683909. Epub 2013 Aug 18. Neural Plast. 2013. PMID: 24024041 Free PMC article. Review.
-
Cholinergic potentiation of visual perception and vision restoration in rodents and humans.Restor Neurol Neurosci. 2019;37(6):553-569. doi: 10.3233/RNN-190947. Restor Neurol Neurosci. 2019. PMID: 31839615 Review.
Cited by
-
Cocaine and Caffeine Effects on the Conditioned Place Preference Test: Concomitant Changes on Early Genes within the Mouse Prefrontal Cortex and Nucleus Accumbens.Front Behav Neurosci. 2017 Oct 18;11:200. doi: 10.3389/fnbeh.2017.00200. eCollection 2017. Front Behav Neurosci. 2017. PMID: 29093669 Free PMC article.
-
Ion channels, guidance molecules, intracellular signaling and transcription factors regulating nervous and vascular system development.J Physiol Sci. 2016 Mar;66(2):175-88. doi: 10.1007/s12576-015-0416-1. Epub 2015 Oct 27. J Physiol Sci. 2016. PMID: 26507418 Free PMC article. Review.
-
Multisensory Integration: Is Medial Prefrontal Cortex Signaling Relevant for the Treatment of Higher-Order Visual Dysfunctions?Front Mol Neurosci. 2022 Jan 17;14:806376. doi: 10.3389/fnmol.2021.806376. eCollection 2021. Front Mol Neurosci. 2022. PMID: 35110996 Free PMC article. Review.
-
Synapse development organized by neuronal activity-regulated immediate-early genes.Exp Mol Med. 2018 Apr 9;50(4):1-7. doi: 10.1038/s12276-018-0025-1. Exp Mol Med. 2018. PMID: 29628504 Free PMC article. Review.
-
H3 and H4 Lysine Acetylation Correlates with Developmental and Experimentally Induced Adult Experience-Dependent Plasticity in the Mouse Visual Cortex.J Exp Neurosci. 2016 Oct 9;10(Suppl 1):49-64. doi: 10.4137/JEN.S39888. eCollection 2016. J Exp Neurosci. 2016. PMID: 27891053 Free PMC article.
References
-
- Amar M, Lucas-Meunier E, Baux G, Fossier P. Blockade of different muscarinic receptor subtypes changes the equilibrium between excitation and inhibition in rat visual cortex. Neuroscience. 2010;169:1610–1620. - PubMed
-
- Baroncelli L, Sale A, Viegi A, Maya Vetencourt JF, De Pasquale R, Baldini S, Maffei L. Experience-dependent reactivation of ocular dominance plasticity in the adult visual cortex. Exp Neurol. 2010;226:100–109. - PubMed
-
- Berardi N, Pizzorusso T, Maffei L. Critical periods during sensory development. Curr Opin Neurobiol. 2000;10:138–145. - PubMed
-
- Cetin A, Komai S, Eliava M, Seeburg PH, Osten P. Stereotaxic gene delivery in the rodent brain. Nat Protoc. 2006;1:3166–3173. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources