

Hepatitis D virus isolates with low replication and epithelial-mesenchymal transition-inducing activity are associated with disease remission
- PMID: 22674995
- PMCID: PMC3416108
- DOI: 10.1128/JVI.00130-12
Hepatitis D virus isolates with low replication and epithelial-mesenchymal transition-inducing activity are associated with disease remission
Abstract
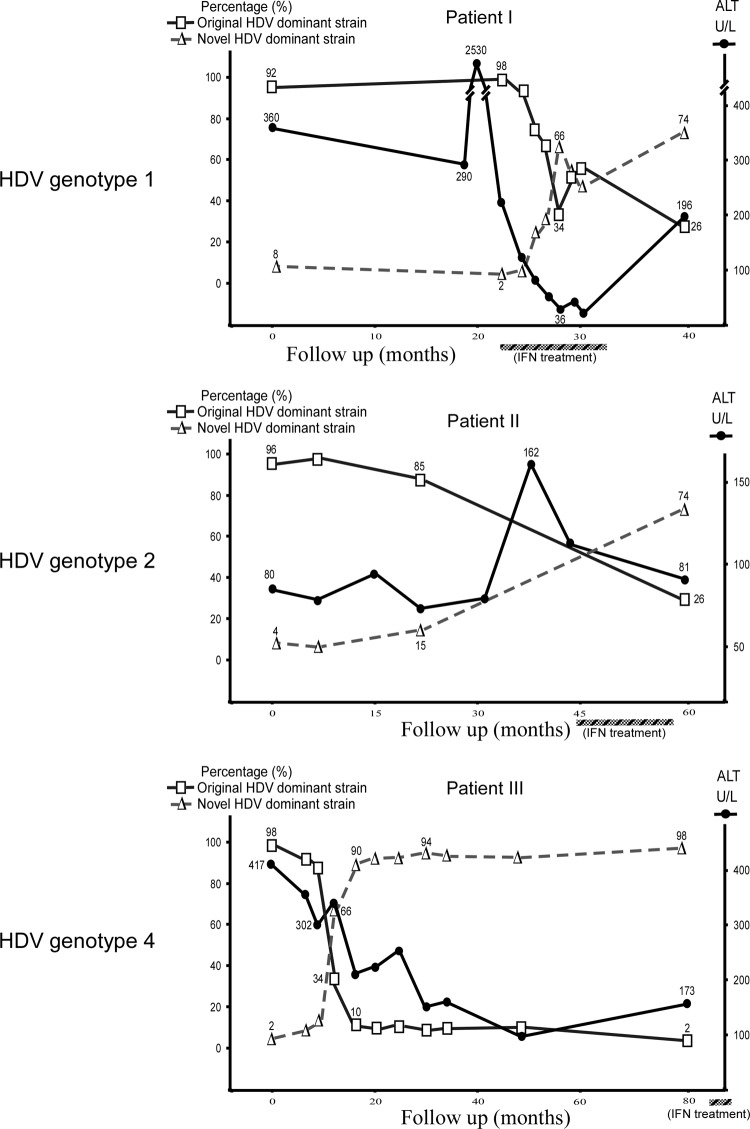
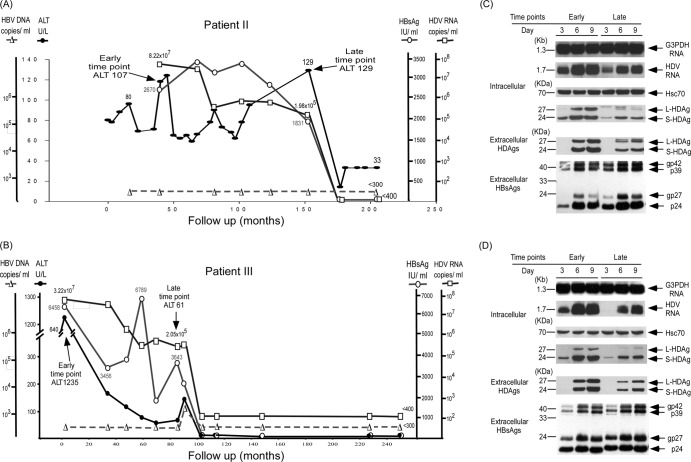
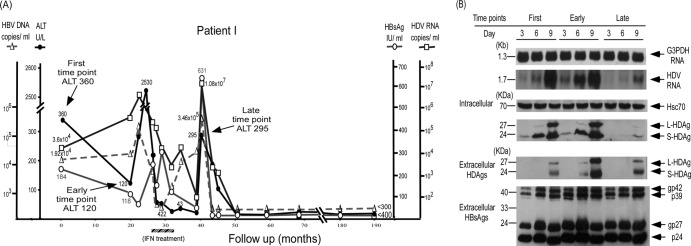
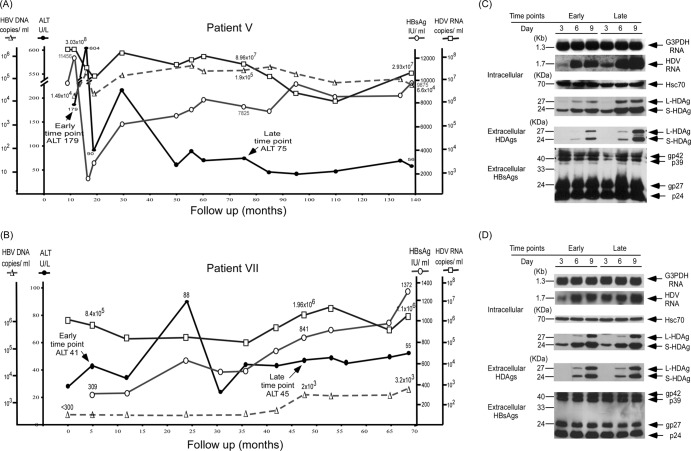
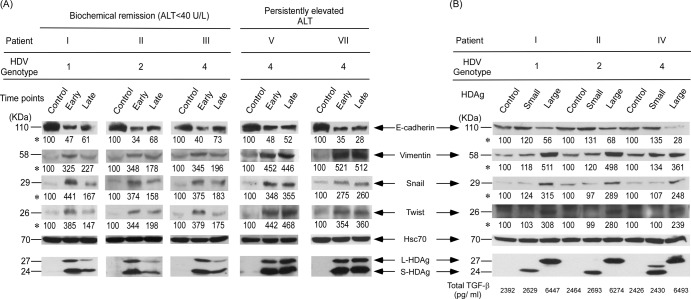
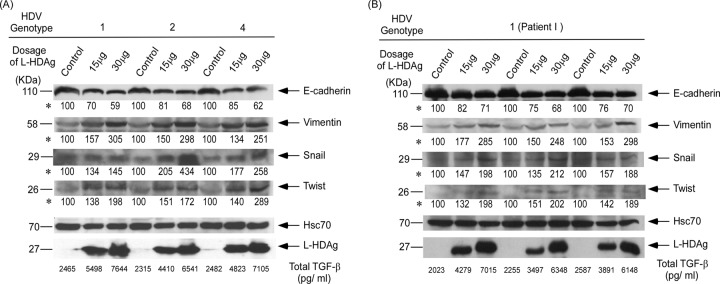
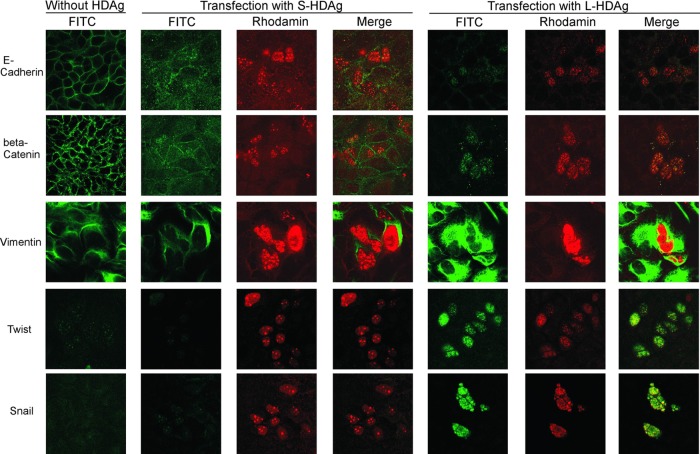
Clearance of hepatitis D virus (HDV) viremia leads to disease remission. Large hepatitis delta antigen (L-HDAg) has been reported to activate transforming growth factor β, which may induce epithelial-mesenchymal transition (EMT) and fibrogenesis. This study analyzed serum HDV RNA "quasispecies" in HDV-infected patients at two stages of infection: before and after alanine aminotransferase (ALT) elevations. Included in the study were four patients who went into remission after ALT elevation and three patients who did not go into remission and progressed to cirrhosis or hepatocellular carcinoma. Full-length HDV cDNA clones were obtained from the most abundant HDV RNA species at the pre- and post-ALT elevation stages. Using an in vitro model consisting of Huh-7 cells transfected with cloned HDV cDNAs, the pre- or post-ALT elevation dominant HDV RNA species were characterized for (i) their replication capacity by measuring HDV RNA and HDAg levels in transfected cells and (ii) their capacity to induce EMT by measuring the levels of the mesenchymal-cell-specific protein vimentin, the EMT regulators twist and snail, and the epithelial-cell-specific protein E-cadherin. Results show that in patients in remission, the post-ALT elevation dominant HDV RNA species had a lower replication capacity in vitro and lower EMT activity than their pre-ALT elevation counterparts. This was not true of patients who did not go into remission. The expression of L-HDAg, but not small HDAg, increased the expression of the EMT-related proteins. It is concluded that in chronically infected patients, HDV quasispecies with a low replication capacity and low EMT activity are associated with disease remission.
Figures
Similar articles
-
Statin inhibits large hepatitis delta antigen-Smad3 -twist-mediated epithelial-to-mesenchymal transition and hepatitis D virus secretion.J Biomed Sci. 2020 May 21;27(1):65. doi: 10.1186/s12929-020-00659-6. J Biomed Sci. 2020. PMID: 32434501 Free PMC article.
-
Genotype-specific complementation of hepatitis delta virus RNA replication by hepatitis delta antigen.J Virol. 1998 Apr;72(4):2806-14. doi: 10.1128/JVI.72.4.2806-2814.1998. J Virol. 1998. PMID: 9525600 Free PMC article.
-
Comprehensive analysis of mutations in the hepatitis delta virus genome based on full-length sequencing in a nationwide cohort study and evolutionary pattern during disease progression.Clin Microbiol Infect. 2015 May;21(5):510.e11-23. doi: 10.1016/j.cmi.2014.12.008. Epub 2014 Dec 26. Clin Microbiol Infect. 2015. PMID: 25656625
-
Functional and clinical significance of hepatitis D virus genotype II infection.Curr Top Microbiol Immunol. 2006;307:173-86. doi: 10.1007/3-540-29802-9_9. Curr Top Microbiol Immunol. 2006. PMID: 16903226 Review.
-
Innate immune recognition and modulation in hepatitis D virus infection.World J Gastroenterol. 2020 Jun 7;26(21):2781-2791. doi: 10.3748/wjg.v26.i21.2781. World J Gastroenterol. 2020. PMID: 32550754 Free PMC article. Review.
Cited by
-
Hepatocellular Carcinoma in Chronic Viral Hepatitis: Where Do We Stand?Int J Mol Sci. 2022 Jan 2;23(1):500. doi: 10.3390/ijms23010500. Int J Mol Sci. 2022. PMID: 35008926 Free PMC article. Review.
-
Hepatitis D and hepatocellular carcinoma.World J Hepatol. 2015 Apr 18;7(5):777-86. doi: 10.4254/wjh.v7.i5.777. World J Hepatol. 2015. PMID: 25914778 Free PMC article. Review.
-
HDV Pathogenesis: Unravelling Ariadne's Thread.Viruses. 2021 Apr 28;13(5):778. doi: 10.3390/v13050778. Viruses. 2021. PMID: 33924806 Free PMC article. Review.
-
Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC.Viruses. 2024 May 21;16(6):817. doi: 10.3390/v16060817. Viruses. 2024. PMID: 38932110 Free PMC article.
-
Life cycle and pathogenesis of hepatitis D virus: A review.World J Hepatol. 2013 Dec 27;5(12):666-75. doi: 10.4254/wjh.v5.i12.666. World J Hepatol. 2013. PMID: 24409335 Free PMC article. Review.
References
-
- Castilla A, Prieto J, Fausto N. 1991. Transforming growth factors beta 1 and alpha in chronic liver disease. Effects of interferon alpha therapy. N. Engl. J. Med. 324:933–940 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
