

The cancer stem cell subtype determines immune infiltration of glioblastoma
- PMID: 22676416
- PMCID: PMC3464079
- DOI: 10.1089/scd.2011.0660
The cancer stem cell subtype determines immune infiltration of glioblastoma
Abstract
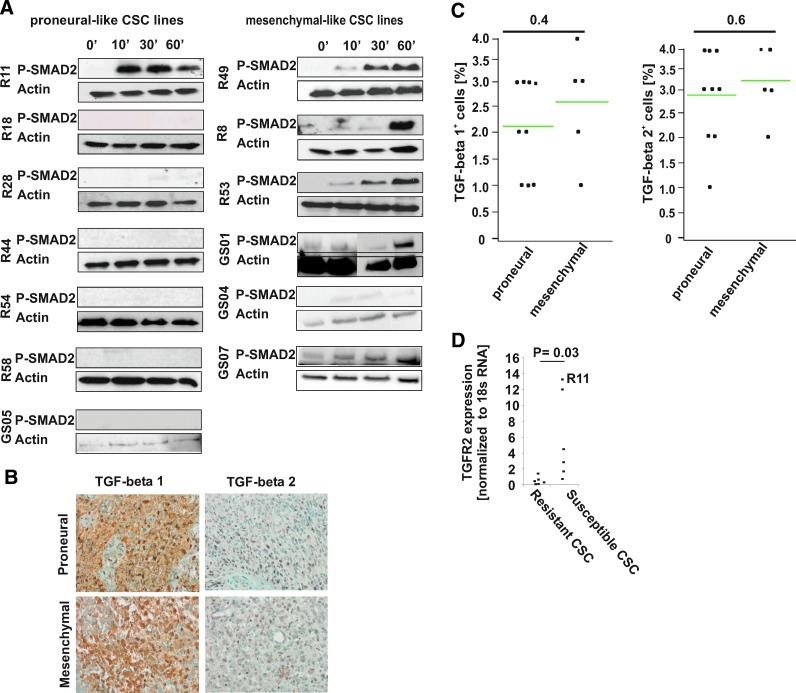
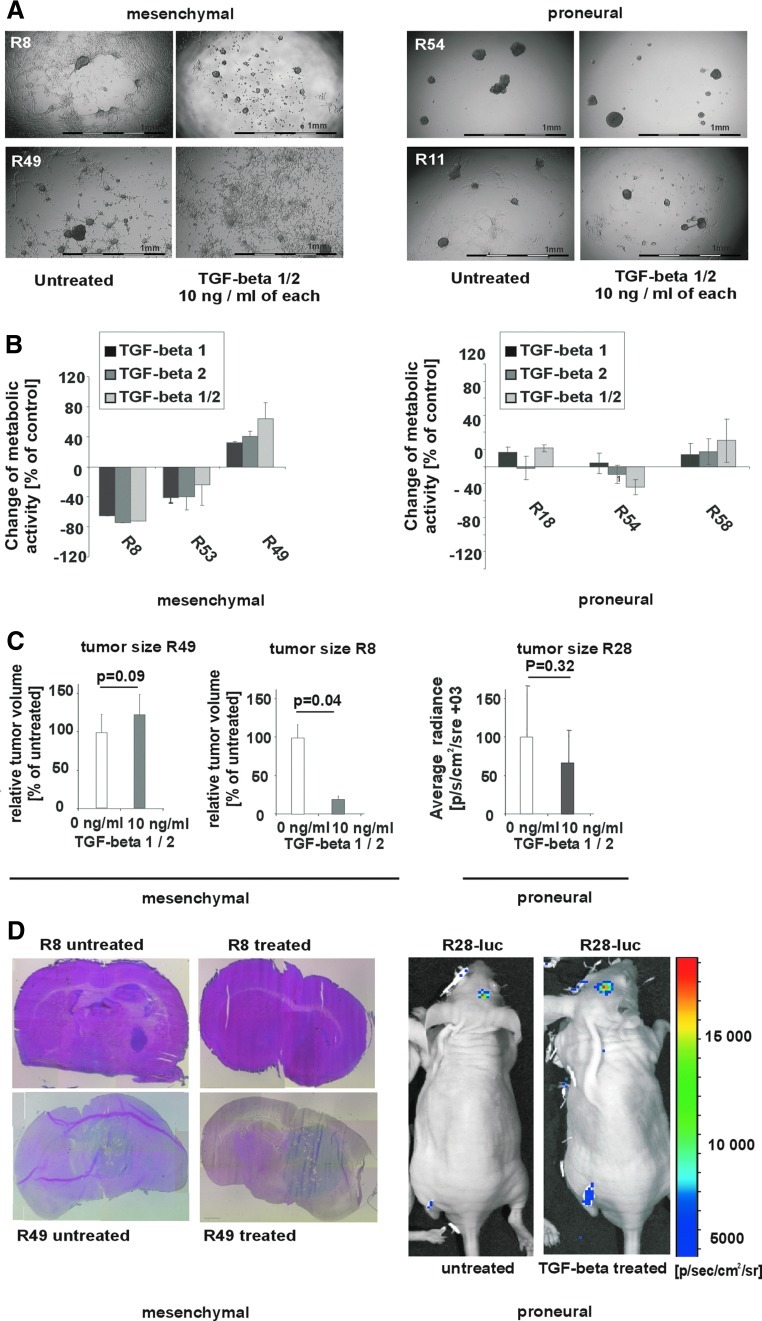
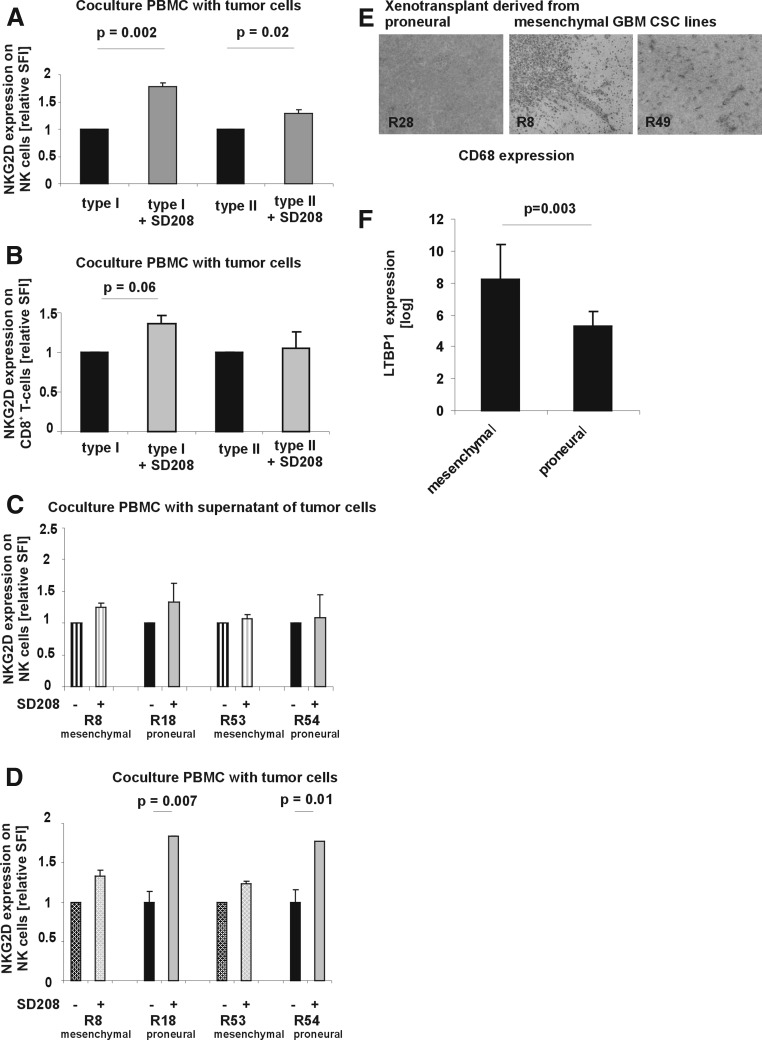
Immune cell infiltration varies widely between different glioblastomas (GBMs). The underlying mechanism, however, remains unknown. Here we show that TGF-beta regulates proliferation, migration, and tumorigenicity of mesenchymal GBM cancer stem cells (CSCs) in vivo and in vitro. In contrast, proneural GBM CSCs resisted TGF-beta due to TGFR2 deficiency. In vivo, a substantially increased infiltration of immune cells was observed in mesenchymal GBMs, while immune infiltrates were rare in proneural GBMs. On a functional level, proneural CSC lines caused a significantly stronger TGF-beta-dependent suppression of NKG2D expression on CD8(+) T and NK cells in vitro providing a mechanistic explanation for the reduced immune infiltration of proneural GBMs. Thus, the molecular subtype of CSCs TGF-beta-dependently contributes to the degree of immune infiltration.
Figures


References
-
- Huse JT. Phillips HS. Brennan CW. Molecular subclassification of diffuse gliomas: seeing order in the chaos. Glia. 2011;59:1190–1199. - PubMed
-
- Phillips HS. Kharbanda S. Chen R. Forrest WF. Soriano RH. Wu TD. Misra A. Nigro JM. Colman H, et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell. 2006;9:157–173. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
