

Bistability, bifurcations, and Waddington's epigenetic landscape
- PMID: 22677291
- PMCID: PMC3372930
- DOI: 10.1016/j.cub.2012.03.045
Bistability, bifurcations, and Waddington's epigenetic landscape
Abstract
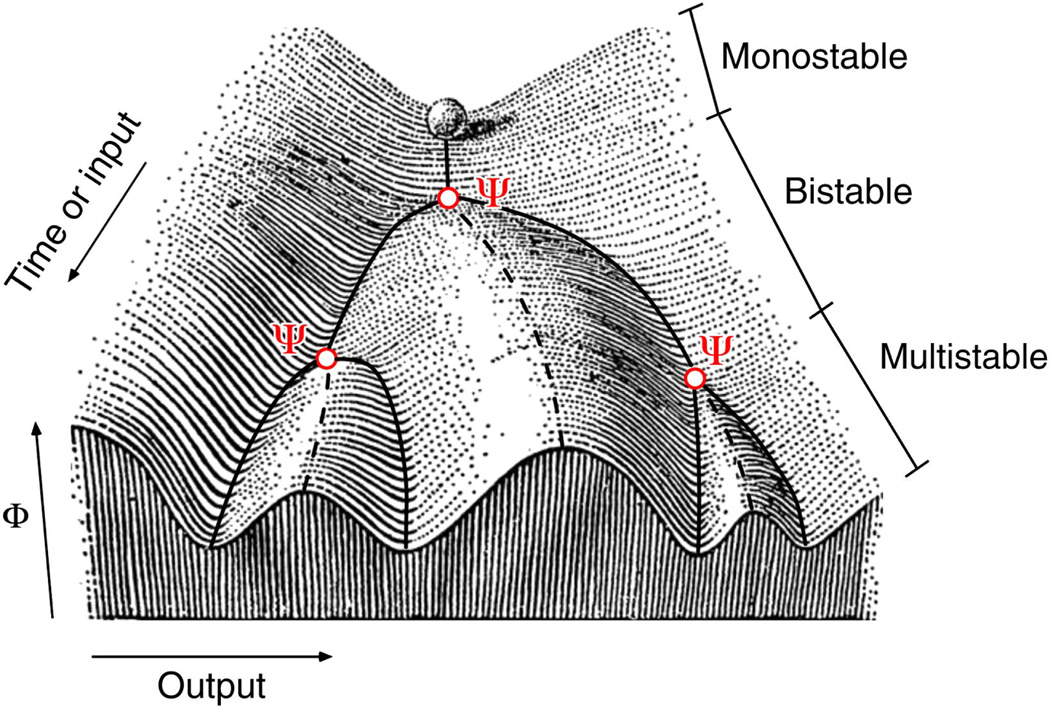
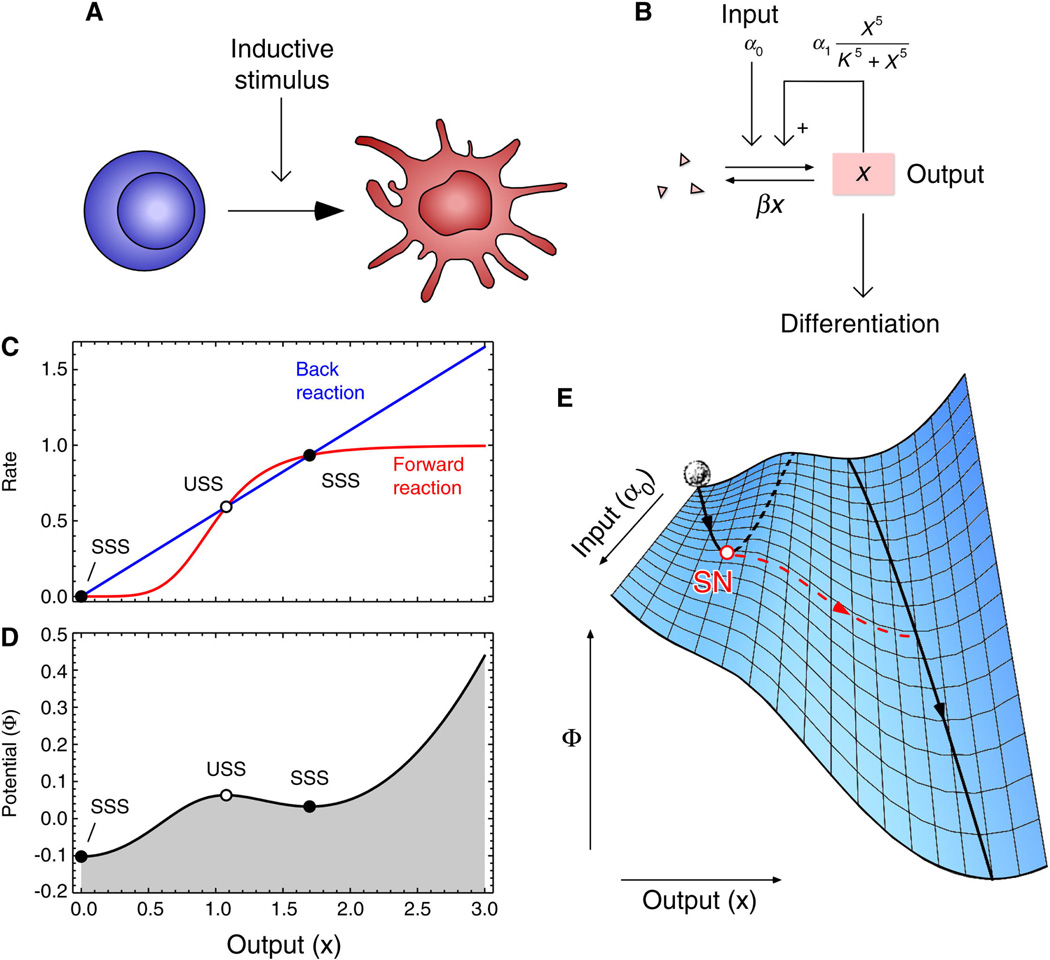
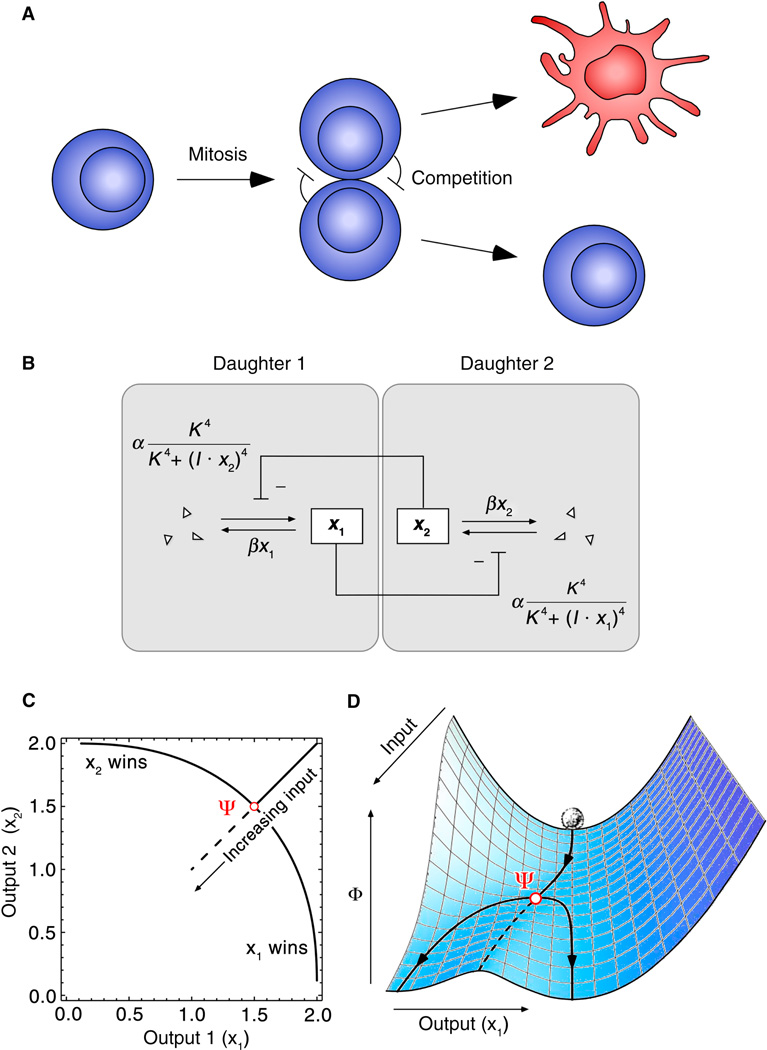
Waddington's epigenetic landscape is probably the most famous and most powerful metaphor in developmental biology. Cells, represented by balls, roll downhill through a landscape of bifurcating valleys. Each new valley represents a possible cell fate and the ridges between the valleys maintain the cell fate once it has been chosen. Here I examine models of two important developmental processes - cell-fate induction and lateral inhibition - and ask whether the landscapes for these models at least qualitatively resemble Waddington's picture. For cell-fate induction, the answer is no. The commitment of a cell to a new fate corresponds to the disappearance of a valley from the landscape, not the splitting of one valley into two, and it occurs through a type of bifurcation - a saddle-node bifurcation - that possesses an intrinsic irreversibility that is missing from Waddington's picture. Lateral inhibition, a symmetrical cell-cell competition process, corresponds better to Waddington's picture, with one valley reversibly splitting into two through a pitchfork bifurcation. I propose an alternative epigenetic landscape that has numerous valleys and ridges right from the start, with the process of cell-fate commitment corresponding to the irreversible disappearance of some of these valleys and ridges, via cell-fate induction, complemented by the creation of new valleys and ridges through processes like cell-cell competition.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Waddington CH. The Strategy of the Genes. London, UK: Volume George Allen and Unwin; 1957.
-
- Strogatz SH. Nonlinear dynamics and chaos: with applications to physics, biology, chemistry, and engineering. Cambridge MA: Westview Press; 1994.
-
- Gilbert SF. Developmental Biology. Ninth Edition. Sunderland MA: Sinauer Associates; 2010.
-
- Delbrück M. Unités Biologiques Douées de Continuité Genetique (international Symposium CNRS No. 8) Paris: Editions du CNRS; 1949. Enzyme systems with alternative steady states; pp. 33–34.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
