

Correlating calcium binding, Förster resonance energy transfer, and conformational change in the biosensor TN-XXL
- PMID: 22677394
- PMCID: PMC3353025
- DOI: 10.1016/j.bpj.2012.03.065
Correlating calcium binding, Förster resonance energy transfer, and conformational change in the biosensor TN-XXL
Abstract
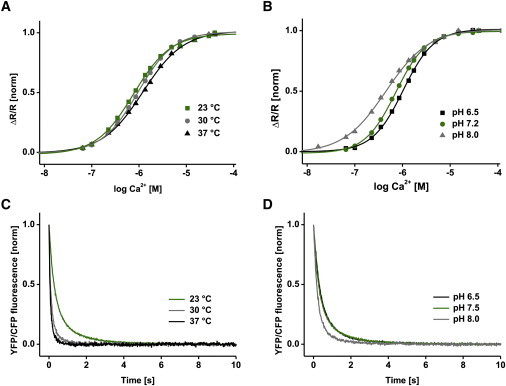
Genetically encoded calcium indicators have become instrumental in imaging signaling in complex tissues and neuronal circuits in vivo. Despite their importance, structure-function relationships of these sensors often remain largely uncharacterized due to their artificial and multimodular composition. Here, we describe a combination of protein engineering and kinetic, spectroscopic, and biophysical analysis of the Förster resonance energy transfer (FRET)-based calcium biosensor TN-XXL. Using fluorescence spectroscopy of engineered tyrosines, we show that two of the four calcium binding EF-hands dominate the FRET output of TN-XXL and that local conformational changes of these hands match the kinetics of FRET change. Using small-angle x-ray scattering and NMR spectroscopy, we show that TN-XXL changes from a flexible elongated to a rigid globular shape upon binding calcium, thus resulting in FRET signal output. Furthermore, we compare calcium titrations using fluorescence lifetime spectroscopy with the ratiometric approach and investigate potential non-FRET effects that may affect the fluorophores. Thus, our data characterize the biophysics of TN-XXL in detail and may form a basis for further rational engineering of FRET-based biosensors.
Copyright © 2012 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Heim R., Tsien R.Y. Engineering green fluorescent protein for improved brightness, longer wavelengths and fluorescence resonance energy transfer. Curr. Biol. 1996;6:178–182. - PubMed
-
- Pollok B.A., Heim R. Using GFP in FRET-based applications. Trends Cell Biol. 1999;9:57–60. - PubMed
-
- Miyawaki A., Tsien R.Y. Monitoring protein conformations and interactions by fluorescence resonance energy transfer between mutants of green fluorescent protein. Methods Enzymol. 2000;327:472–500. - PubMed
-
- Yasuda R. Imaging spatiotemporal dynamics of neuronal signaling using fluorescence resonance energy transfer and fluorescence lifetime imaging microscopy. Curr. Opin. Neurobiol. 2006;16:551–561. - PubMed
-
- Miyawaki A. Development of probes for cellular functions using fluorescent proteins and fluorescence resonance energy transfer. Annu. Rev. Biochem. 2011;80:357–373. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
