

Alzheimer's disease, oestrogen and mitochondria: an ambiguous relationship
- PMID: 22678467
- PMCID: PMC3443477
- DOI: 10.1007/s12035-012-8281-x
Alzheimer's disease, oestrogen and mitochondria: an ambiguous relationship
Abstract
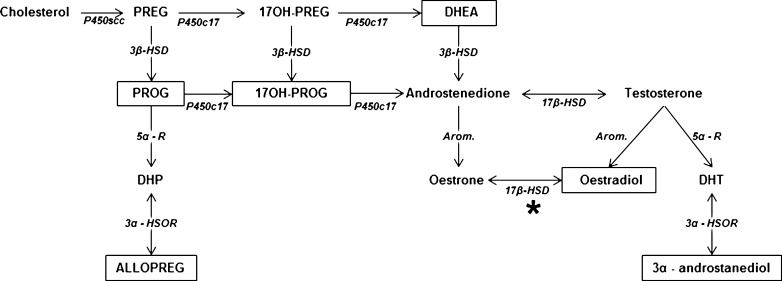
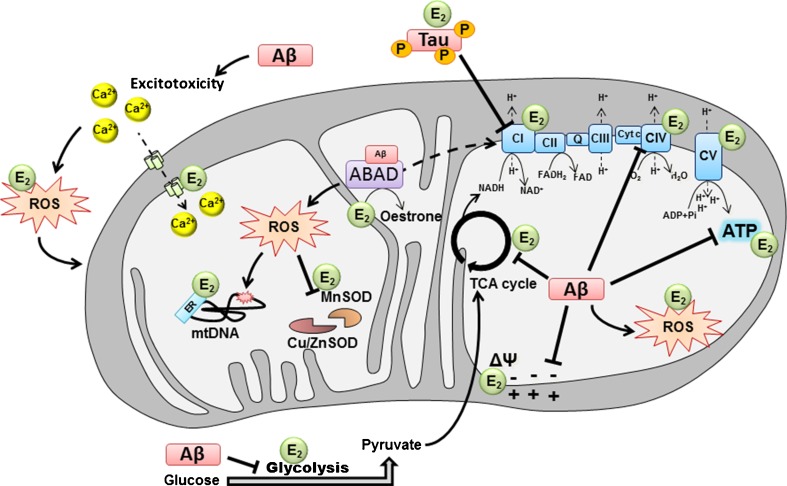
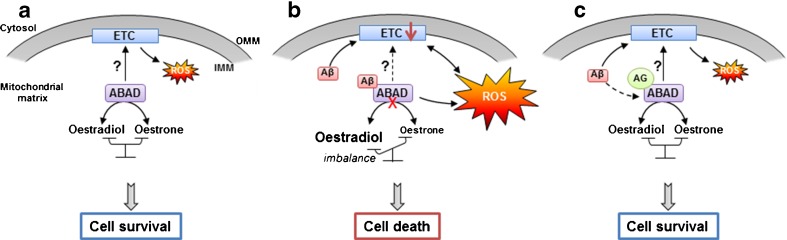
Hormonal deficit in post-menopausal women has been proposed to be one risk factor in Alzheimer's disease (AD) since two thirds of AD patients are women. However, large treatment trials showed negative effects of long-term treatment with oestrogens in older women. Thus, oestrogen treatment after menopause is still under debate, and several hypotheses trying to explain the failure in outcome are under discussion. Concurrently, it was shown that amyloid-beta (Aβ) peptide, the main constituent of senile plaques, as well as abnormally hyperphosphorylated tau protein, the main component of neurofibrillary tangles, can modulate the level of neurosteroids which notably represent neuroactive steroids synthetized within the nervous system, independently of peripheral endocrine glands. In this review, we summarize the role of neurosteroids especially that of oestrogen in AD and discuss their potentially neuroprotective effects with specific regard to the role of oestrogens on the maintenance and function of mitochondria, important organelles which are highly vulnerable to Aβ- and tau-induced toxicity. We also discuss the role of Aβ-binding alcohol dehydrogenase (ABAD), a mitochondrial enzyme able to bind Aβ peptide thereby modifying mitochondrial function as well as oestradiol levels suggesting possible modes of interaction between the three, and the potential therapeutic implication of inhibiting Aβ-ABAD interaction.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Mensah-Nyagan AG, Do-Rego JL, Beaujean D, Luu-The V, Pelletier G, Vaudry H. Neurosteroids: expression of steroidogenic enzymes and regulation of steroid biosynthesis in the central nervous system. Pharmacol Rev. 1999;51(1):63–81. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
