

Actin network architecture can determine myosin motor activity
- PMID: 22679097
- PMCID: PMC3649007
- DOI: 10.1126/science.1221708
Actin network architecture can determine myosin motor activity
Abstract
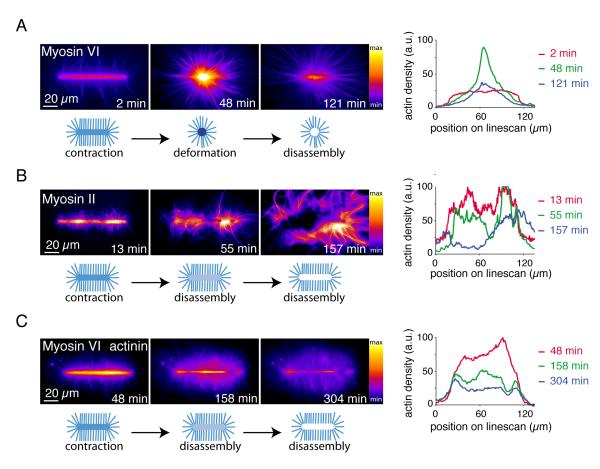
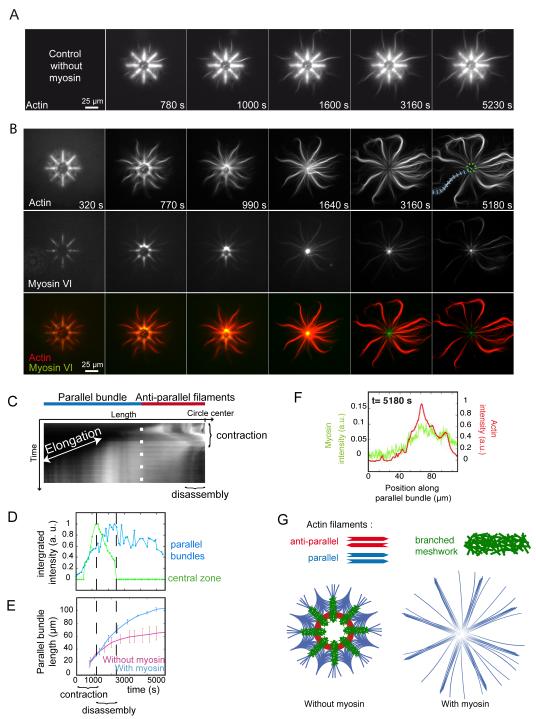
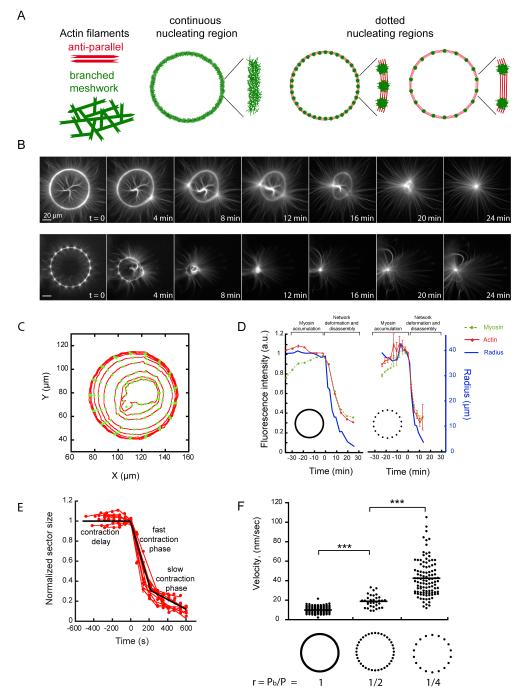
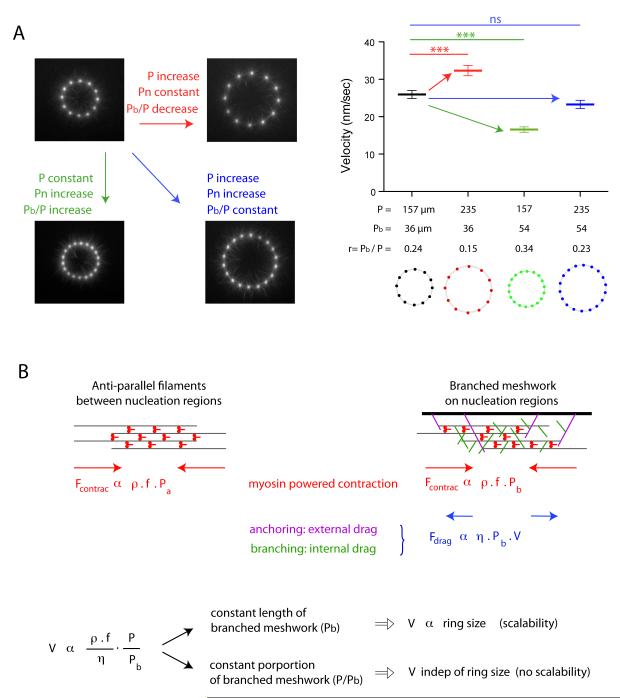
The organization of actin filaments into higher-ordered structures governs eukaryotic cell shape and movement. Global actin network size and architecture are maintained in a dynamic steady state through regulated assembly and disassembly. Here, we used experimentally defined actin structures in vitro to investigate how the activity of myosin motors depends on network architecture. Direct visualization of filaments revealed myosin-induced actin network deformation. During this reorganization, myosins selectively contracted and disassembled antiparallel actin structures, while parallel actin bundles remained unaffected. The local distribution of nucleation sites and the resulting orientation of actin filaments appeared to regulate the scalability of the contraction process. This "orientation selection" mechanism for selective contraction and disassembly suggests how the dynamics of the cellular actin cytoskeleton can be spatially controlled by actomyosin contractility.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
