

The circadian neuropeptide PDF signals preferentially through a specific adenylate cyclase isoform AC3 in M pacemakers of Drosophila
- PMID: 22679392
- PMCID: PMC3367976
- DOI: 10.1371/journal.pbio.1001337
The circadian neuropeptide PDF signals preferentially through a specific adenylate cyclase isoform AC3 in M pacemakers of Drosophila
Abstract
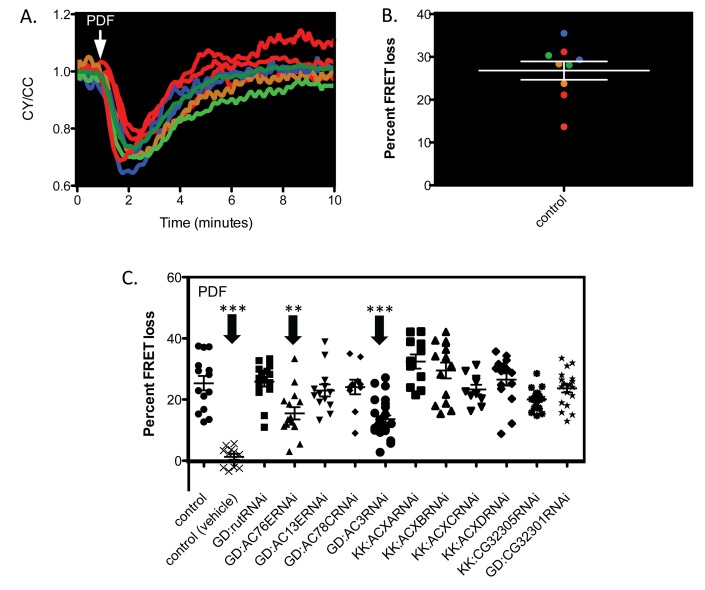
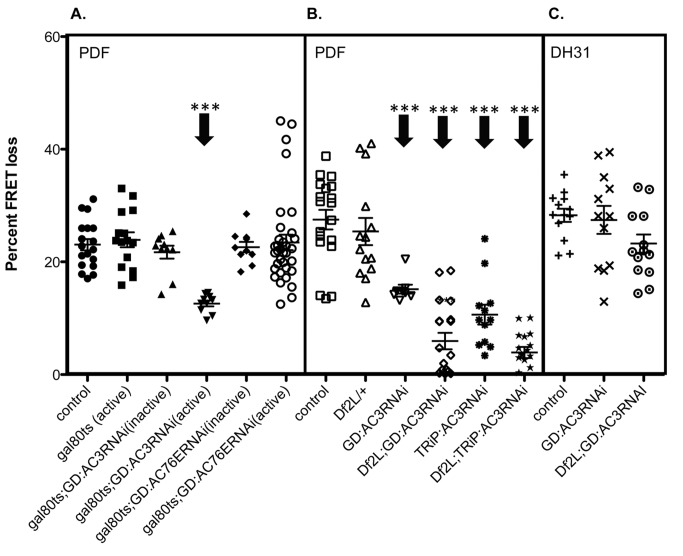
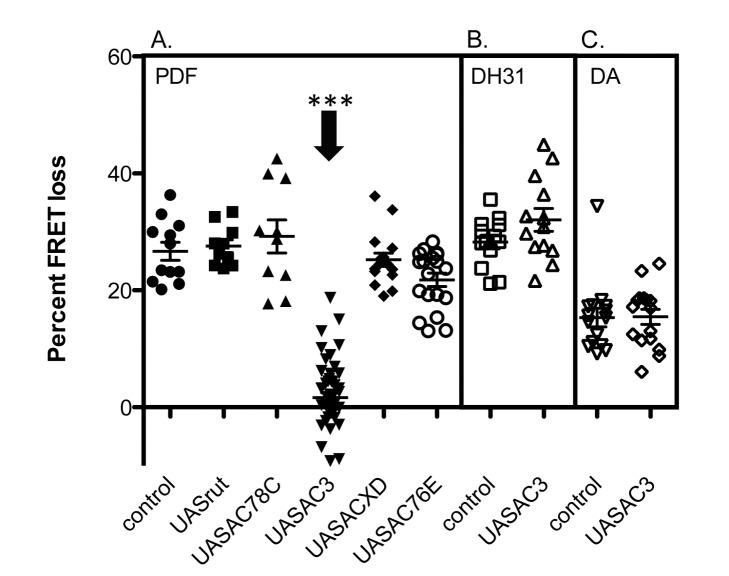
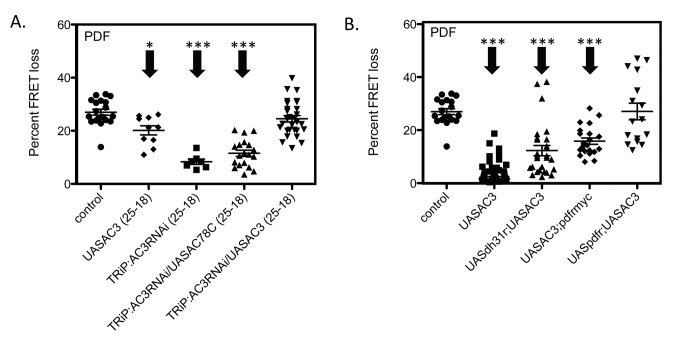
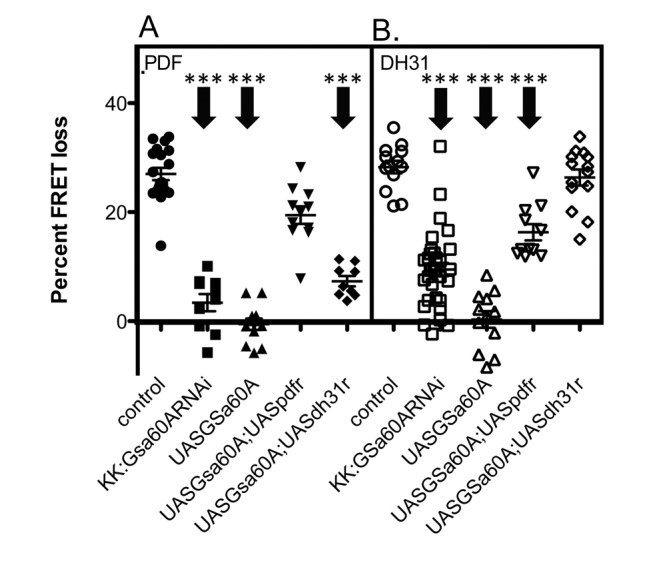
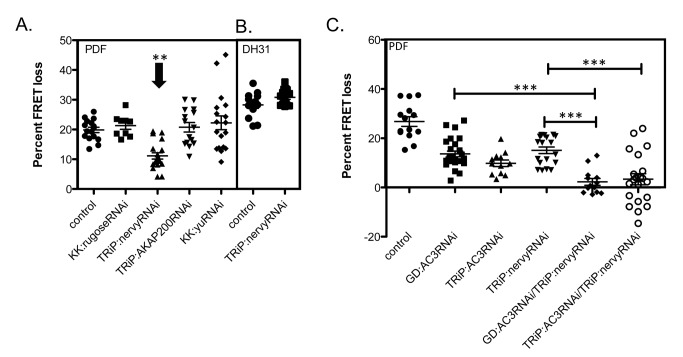
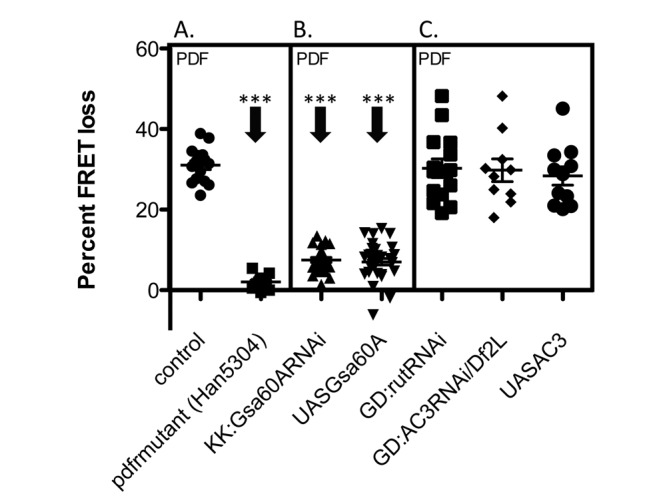
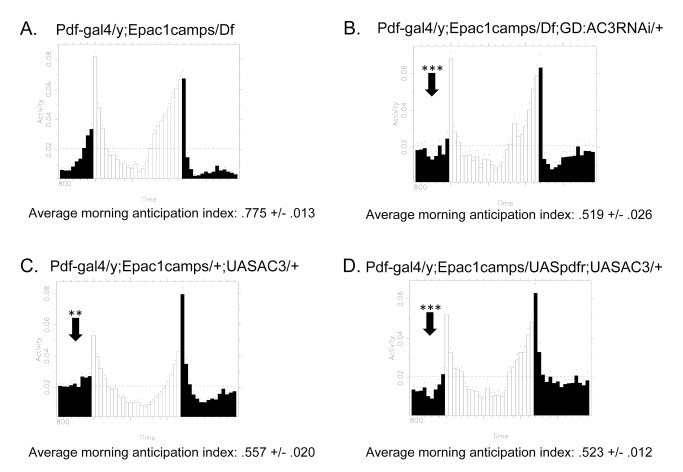
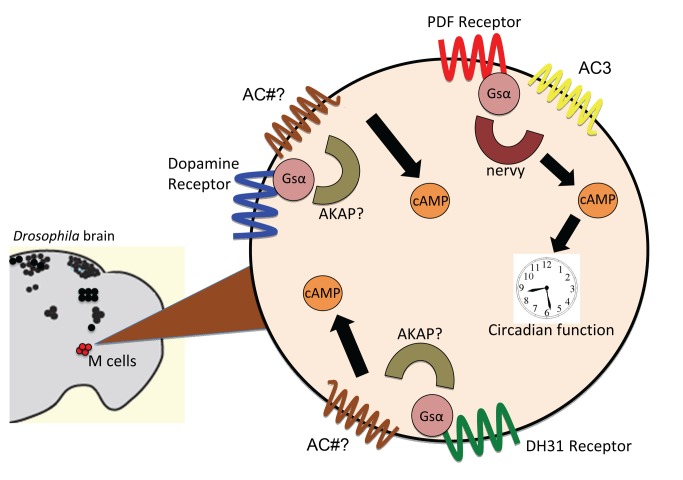
The neuropeptide Pigment Dispersing Factor (PDF) is essential for normal circadian function in Drosophila. It synchronizes the phases of M pacemakers, while in E pacemakers it decelerates their cycling and supports their amplitude. The PDF receptor (PDF-R) is present in both M and subsets of E cells. Activation of PDF-R stimulates cAMP increases in vitro and in M cells in vivo. The present study asks: What is the identity of downstream signaling components that are associated with PDF receptor in specific circadian pacemaker neurons? Using live imaging of intact fly brains and transgenic RNAi, we show that adenylate cyclase AC3 underlies PDF signaling in M cells. Genetic disruptions of AC3 specifically disrupt PDF responses: they do not affect other Gs-coupled GPCR signaling in M cells, they can be rescued, and they do not represent developmental alterations. Knockdown of the Drosophila AKAP-like scaffolding protein Nervy also reduces PDF responses. Flies with AC3 alterations show behavioral syndromes consistent with known roles of M pacemakers as mediated by PDF. Surprisingly, disruption of AC3 does not alter PDF responses in E cells--the PDF-R(+) LNd. Within M pacemakers, PDF-R couples preferentially to a single AC, but PDF-R association with a different AC(s) is needed to explain PDF signaling in the E pacemakers. Thus critical pathways of circadian synchronization are mediated by highly specific second messenger components. These findings support a hypothesis that PDF signaling components within target cells are sequestered into "circadian signalosomes," whose compositions differ between E and M pacemaker cell types.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
E and M circadian pacemaker neurons use different PDF receptor signalosome components in drosophila.J Biol Rhythms. 2013 Aug;28(4):239-48. doi: 10.1177/0748730413497179. J Biol Rhythms. 2013. PMID: 23929551 Free PMC article.
-
Signaling of pigment-dispersing factor (PDF) in the Madeira cockroach Rhyparobia maderae.PLoS One. 2014 Sep 30;9(9):e108757. doi: 10.1371/journal.pone.0108757. eCollection 2014. PLoS One. 2014. PMID: 25269074 Free PMC article.
-
The Drosophila Receptor Protein Tyrosine Phosphatase LAR Is Required for Development of Circadian Pacemaker Neuron Processes That Support Rhythmic Activity in Constant Darkness But Not during Light/Dark Cycles.J Neurosci. 2016 Mar 30;36(13):3860-70. doi: 10.1523/JNEUROSCI.4523-15.2016. J Neurosci. 2016. PMID: 27030770 Free PMC article.
-
Circadian pathway: the other shoe drops.Curr Biol. 2005 Dec 20;15(24):R987-9. doi: 10.1016/j.cub.2005.11.053. Curr Biol. 2005. PMID: 16360675 Review.
-
Mechanisms of clock output in the Drosophila circadian pacemaker system.J Biol Rhythms. 2006 Dec;21(6):445-57. doi: 10.1177/0748730406293910. J Biol Rhythms. 2006. PMID: 17107935 Review.
Cited by
-
Mating disrupts morning anticipation in Drosophila melanogaster females.PLoS Genet. 2022 Dec 22;18(12):e1010258. doi: 10.1371/journal.pgen.1010258. eCollection 2022 Dec. PLoS Genet. 2022. PMID: 36548223 Free PMC article.
-
Discovery of levodopa-induced dyskinesia-associated genes using genomic studies in patients and Drosophila behavioral analyses.Commun Biol. 2022 Aug 25;5(1):872. doi: 10.1038/s42003-022-03830-x. Commun Biol. 2022. PMID: 36008531 Free PMC article.
-
Circadian regulation of the Na+/K+-ATPase alpha subunit in the visual system is mediated by the pacemaker and by retina photoreceptors in Drosophila melanogaster.PLoS One. 2013 Sep 10;8(9):e73690. doi: 10.1371/journal.pone.0073690. eCollection 2013. PLoS One. 2013. PMID: 24040028 Free PMC article.
-
Enhanced sleep reverses memory deficits and underlying pathology in Drosophila models of Alzheimer's disease.Neurobiol Sleep Circadian Rhythms. 2017 Jan;2:15-26. doi: 10.1016/j.nbscr.2016.09.001. Epub 2016 Sep 28. Neurobiol Sleep Circadian Rhythms. 2017. PMID: 29094110 Free PMC article.
-
Regulation of PDF receptor signaling controlling daily locomotor rhythms in Drosophila.PLoS Genet. 2022 May 23;18(5):e1010013. doi: 10.1371/journal.pgen.1010013. eCollection 2022 May. PLoS Genet. 2022. PMID: 35605015 Free PMC article.
References
-
- Nitabach M. N, Taghert P. H. Organization of the Drosophila circadian control circuit. Curr Bio. 2008;18(2):R84–R93. - PubMed
-
- Grima B, Chelot E, Xia R, Rouyer F. Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature. 2004;431:869–873. - PubMed
-
- Stoleru D, Peng Y, Agosto J, Rosbash M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature. 2004;431:862–868. - PubMed
-
- Yoshii T, Funada Y, Ibuki-Ishibashi T, Matsumoto A, Tanimura T, Tomioka K. Drosophila cryb mutation reveals two circadian clocks that drive locomotor rhythm and have different responsiveness to light. J Insect Physiol. 2004;50(6):479–488. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
