

The SNARE Machinery in Mast Cell Secretion
- PMID: 22679448
- PMCID: PMC3367400
- DOI: 10.3389/fimmu.2012.00143
The SNARE Machinery in Mast Cell Secretion
Abstract
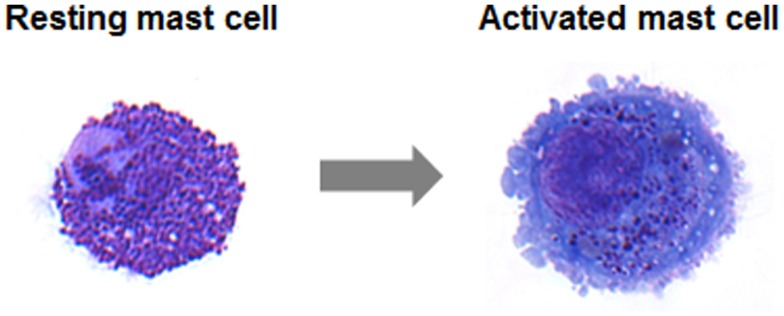
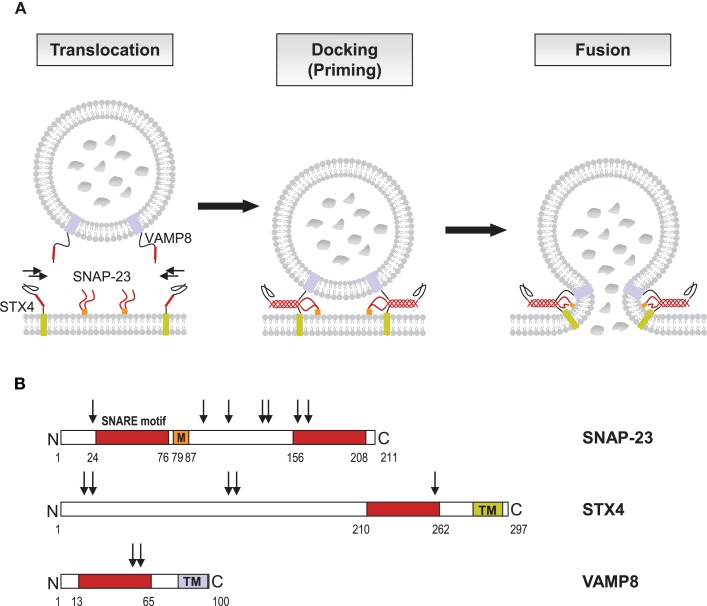
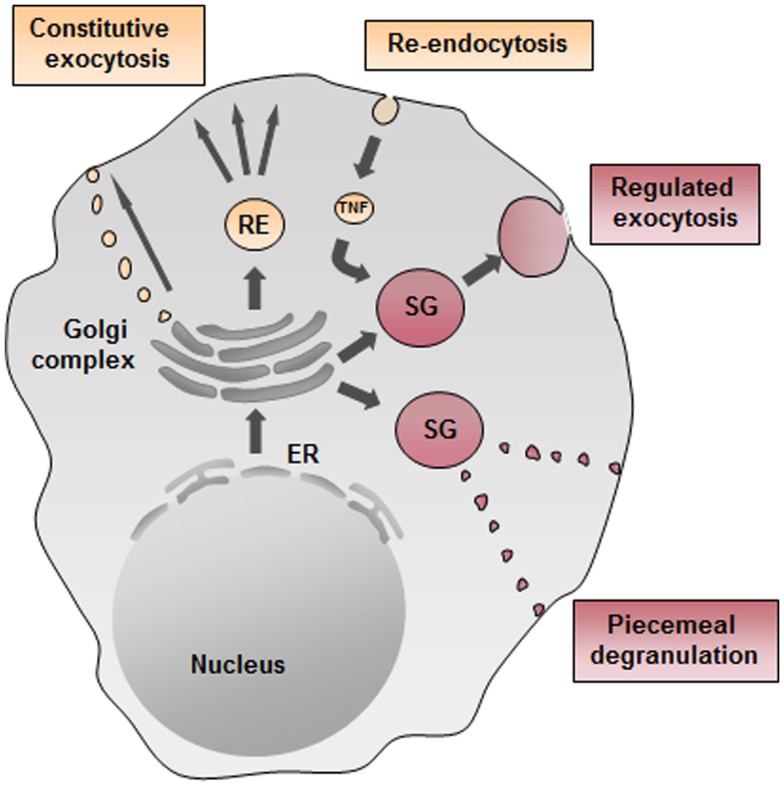
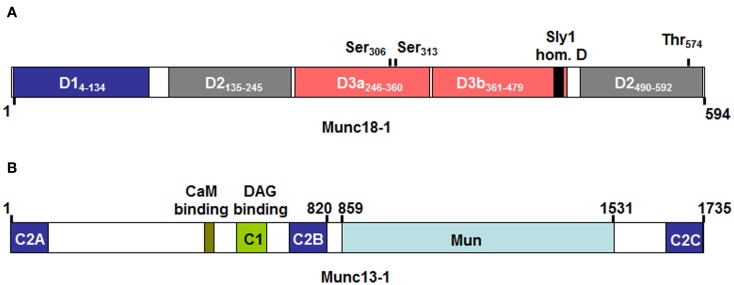
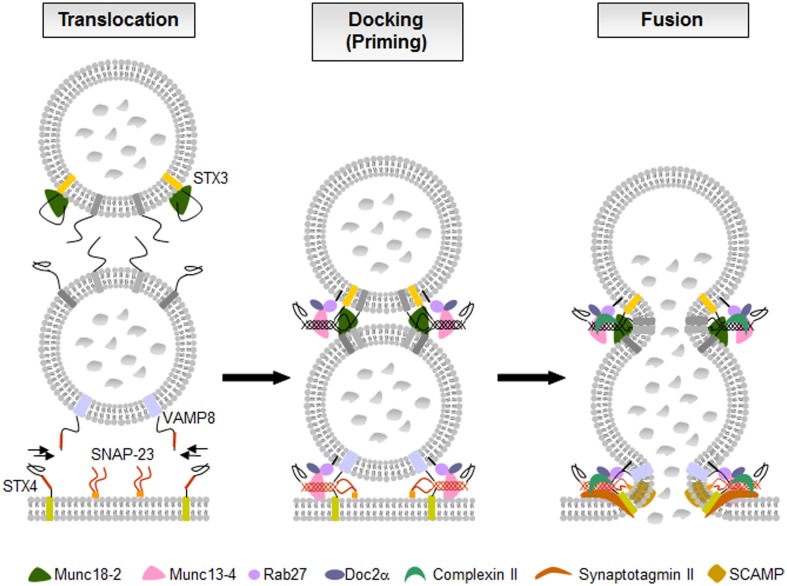
Mast cells are known as inflammatory cells which exert their functions in allergic and anaphylactic reactions by secretion of numerous inflammatory mediators. During an allergic response, the high-affinity IgE receptor, FcεRI, becomes cross-linked by receptor-bound IgE and antigen resulting in immediate release of pre-synthesized mediators - stored in granules - as well as in de novo synthesis of various mediators like cytokines and chemokines. Soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP) receptors (SNARE) proteins were found to play a central role in regulating membrane fusion events during exocytosis. In addition, several accessory regulators like Munc13, Munc18, Rab GTPases, secretory carrier membrane proteins, complexins, or synaptotagmins were found to be involved in membrane fusion. In this review we summarize our current knowledge about the SNARE machinery and its mechanism of action in mast cell secretion.
Keywords: SNARE proteins; exocytosis; mast cell.
Figures
Similar articles
-
The mechanisms of exocytosis in mast cells.Adv Exp Med Biol. 2011;716:107-22. doi: 10.1007/978-1-4419-9533-9_7. Adv Exp Med Biol. 2011. PMID: 21713654 Review.
-
Soluble NSF attachment protein receptors (SNAREs) in RBL-2H3 mast cells: functional role of syntaxin 4 in exocytosis and identification of a vesicle-associated membrane protein 8-containing secretory compartment.J Immunol. 2000 Jun 1;164(11):5850-7. doi: 10.4049/jimmunol.164.11.5850. J Immunol. 2000. PMID: 10820264
-
SNAP-23 and syntaxin-3 are required for chemokine release by mature human mast cells.Mol Immunol. 2011 Oct;49(1-2):353-8. doi: 10.1016/j.molimm.2011.09.011. Epub 2011 Oct 6. Mol Immunol. 2011. PMID: 21981832
-
Mast cell degranulation requires N-ethylmaleimide-sensitive factor-mediated SNARE disassembly.J Immunol. 2003 Nov 15;171(10):5345-52. doi: 10.4049/jimmunol.171.10.5345. J Immunol. 2003. PMID: 14607937
-
SNAREs and associated regulators in the control of exocytosis in the RBL-2H3 mast cell line.Mol Immunol. 2002 Sep;38(16-18):1341-5. doi: 10.1016/s0161-5890(02)00085-8. Mol Immunol. 2002. PMID: 12217405 Review.
Cited by
-
Anaphylactic Degranulation of Mast Cells: Focus on Compound Exocytosis.J Immunol Res. 2019 Mar 18;2019:9542656. doi: 10.1155/2019/9542656. eCollection 2019. J Immunol Res. 2019. PMID: 31011586 Free PMC article. Review.
-
Soluble N-Ethylmaleimide-Sensitive Factor Attachment Protein Receptor-Derived Peptides for Regulation of Mast Cell Degranulation.Front Immunol. 2018 Apr 11;9:725. doi: 10.3389/fimmu.2018.00725. eCollection 2018. Front Immunol. 2018. PMID: 29696021 Free PMC article.
-
Kinesin-1 controls mast cell degranulation and anaphylaxis through PI3K-dependent recruitment to the granular Slp3/Rab27b complex.J Cell Biol. 2016 Oct 24;215(2):203-216. doi: 10.1083/jcb.201605073. J Cell Biol. 2016. PMID: 27810912 Free PMC article.
-
The Circadian Clock Drives Mast Cell Functions in Allergic Reactions.Front Immunol. 2018 Jul 6;9:1526. doi: 10.3389/fimmu.2018.01526. eCollection 2018. Front Immunol. 2018. PMID: 30034393 Free PMC article. Review.
-
Activation of P2X7 Receptors in Peritoneal and Meningeal Mast Cells Detected by Uptake of Organic Dyes: Possible Purinergic Triggers of Neuroinflammation in Meninges.Front Cell Neurosci. 2019 Feb 13;13:45. doi: 10.3389/fncel.2019.00045. eCollection 2019. Front Cell Neurosci. 2019. PMID: 30814932 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources