

Ketoglutarate transport protein KgtP is secreted through the type III secretion system and contributes to virulence in Xanthomonas oryzae pv. oryzae
- PMID: 22685129
- PMCID: PMC3406143
- DOI: 10.1128/AEM.07997-11
Ketoglutarate transport protein KgtP is secreted through the type III secretion system and contributes to virulence in Xanthomonas oryzae pv. oryzae
Abstract
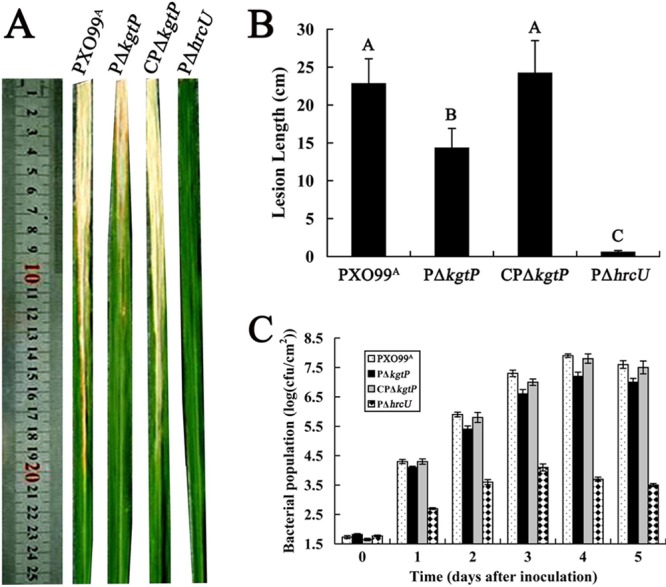
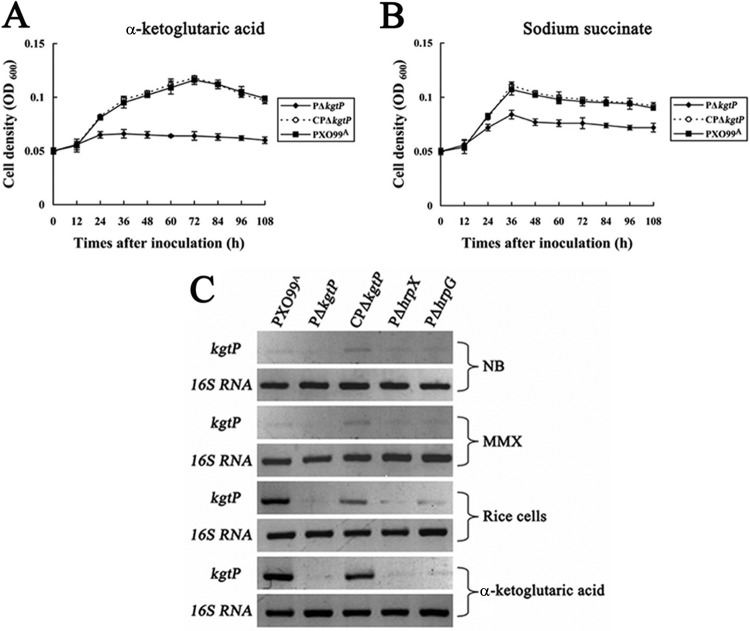
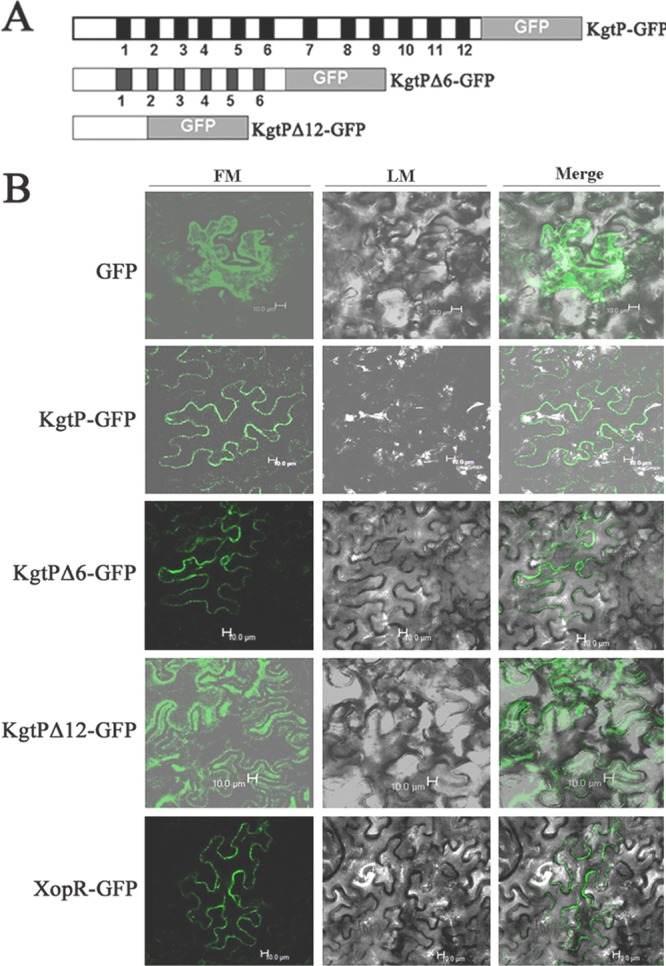
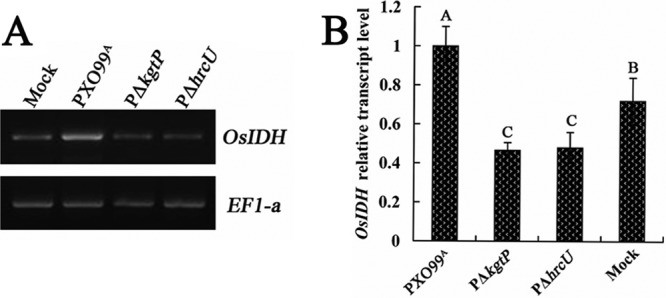
The phytopathogenic prokaryote Xanthomonas oryzae pv. oryzae is the causal agent of bacterial leaf blight (BB) of rice and utilizes a type III secretion system (T3SS) to deliver T3SS effectors into rice cells. In this report, we show that the ketoglutarate transport protein (KgtP) is secreted in an HpaB-independent manner through the T3SS of X. oryzae pv. oryzae PXO99(A) and localizes to the host cell membrane for α-ketoglutaric acid export. kgtP contained an imperfect PIP box (plant-inducible promoter) in the promoter region and was positively regulated by HrpX and HrpG. A kgtP deletion mutant was impaired in bacterial virulence and growth in planta; furthermore, the mutant showed reduced growth in minimal media containing α-ketoglutaric acid or sodium succinate as the sole carbon source. The reduced virulence and the deficiency in α-ketoglutaric acid utilization by the kgtP mutant were restored to wild-type levels by the presence of kgtP in trans. The expression of OsIDH, which is responsible for the synthesis of α-ketoglutaric acid in rice, was enhanced when KgtP was present in the pathogen. To our knowledge, this is the first report demonstrating that KgtP, which is regulated by HrpG and HrpX and secreted by the T3SS in Xanthomonas oryzae pv. oryzae, transports α-ketoglutaric acid when the pathogen infects rice.
Figures
References
-
- Akimoto-Tomiyama C, et al. 2012. XopR, a type III effector secreted by Xanthomonas oryzae pv. oryzae, suppresses microbe-associated molecular pattern-triggered immunity in Arabidopsis thaliana. Mol. Plant Microbe Interact. 25:505–514 - PubMed
-
- Alfano JR, Collmer A. 2004. Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annu. Rev. Phytopathol. 42:385–414 - PubMed
-
- Arwas R, McKay IA, Rowney FRP, Dilworth MJ, Glenn AR. 1985. Properties of organic acid utilization mutants of Rhizobium leguminosarum strain 300. J. Gen. Microbiol. 131:2059–2066
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
