

Anoctamin 1 (Tmem16A) Ca2+-activated chloride channel stoichiometrically interacts with an ezrin-radixin-moesin network
- PMID: 22685202
- PMCID: PMC3387097
- DOI: 10.1073/pnas.1200174109
Anoctamin 1 (Tmem16A) Ca2+-activated chloride channel stoichiometrically interacts with an ezrin-radixin-moesin network
Abstract
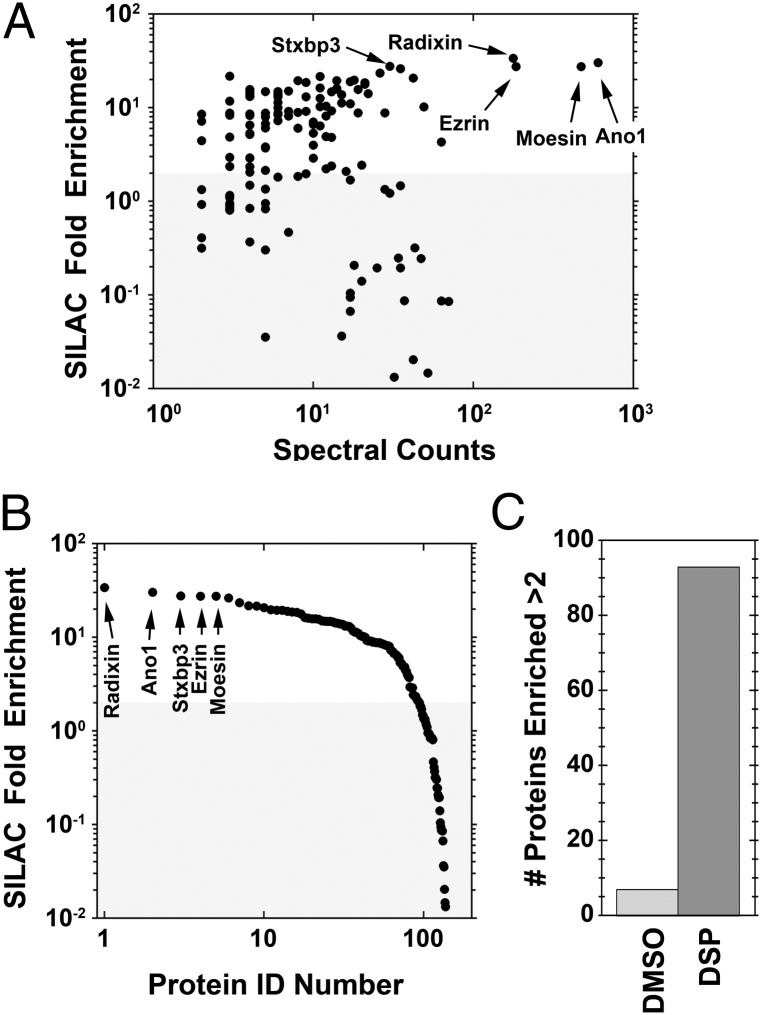
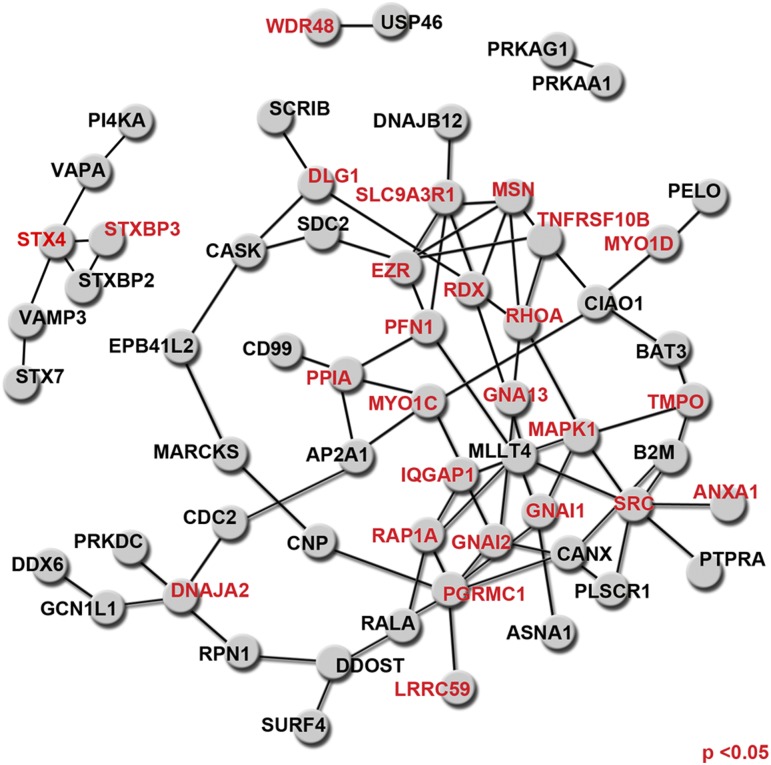
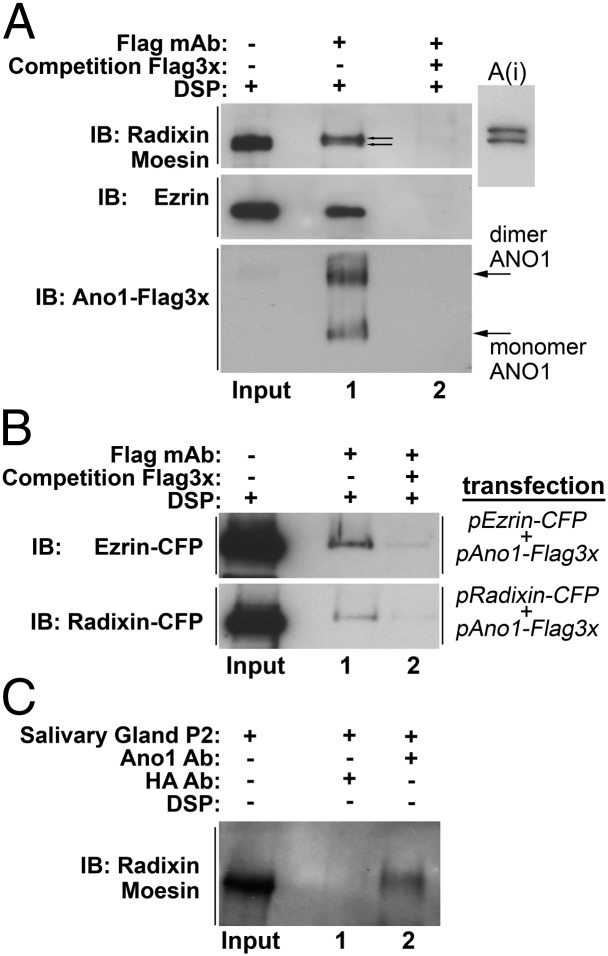
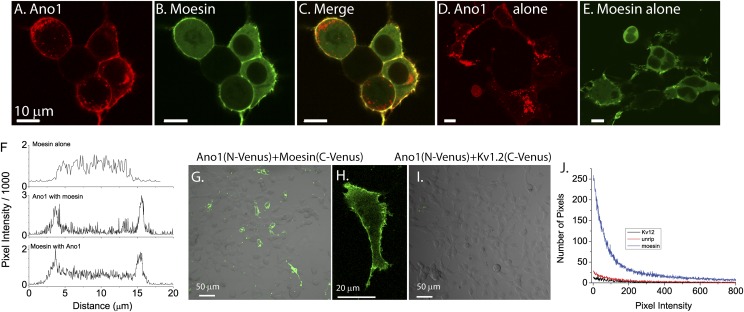
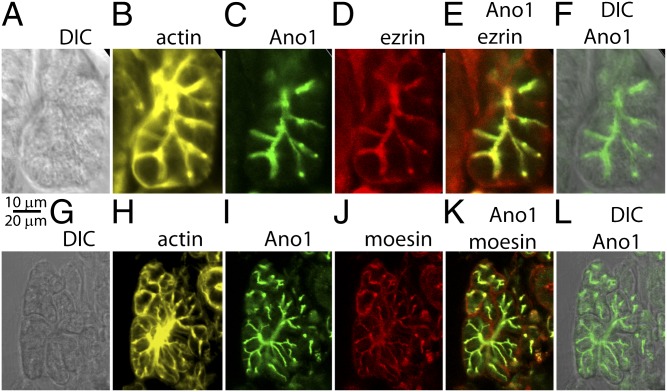
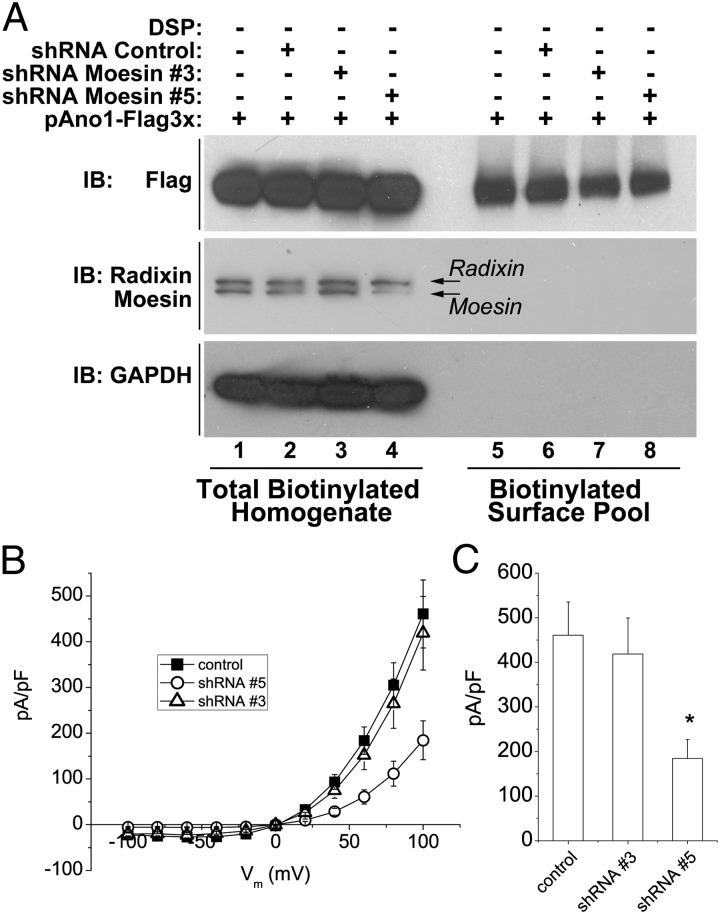
The newly discovered Ca(2+)-activated Cl(-) channel (CaCC), Anoctamin 1 (Ano1 or TMEM16A), has been implicated in vital physiological functions including epithelial fluid secretion, gut motility, and smooth muscle tone. Overexpression of Ano1 in HEK cells or Xenopus oocytes is sufficient to generate Ca(2+)-activated Cl(-) currents, but the details of channel composition and the regulatory factors that control channel biology are incompletely understood. We used a highly sensitive quantitative SILAC proteomics approach to obtain insights into stoichiometric protein networks associated with the Ano1 channel. These studies provide a comprehensive footprint of putative Ano1 regulatory networks. We find that Ano1 associates with the signaling/scaffolding proteins ezrin, radixin, moesin, and RhoA, which link the plasma membrane to the cytoskeleton with very high stoichiometry. Ano1, ezrin, and moesin/radixin colocalize apically in salivary gland epithelial cells, and overexpression of moesin and Ano1 in HEK cells alters the subcellular localization of both proteins. Moreover, interfering RNA for moesin modifies Ano1 current without affecting its surface expression level. Another network associated with Ano1 includes the SNARE and SM proteins VAMP3, syntaxins 2 and -4, and syntaxin-binding proteins munc18b and munc18c, which are integral to translocation of vesicles to the plasma membrane. A number of other regulatory proteins, including GTPases, Ca(2+)-binding proteins, kinases, and lipid-interacting proteins are enriched in the Ano1 complex. These data provide stoichiometrically prioritized information about mechanisms regulating Ano1 function and trafficking to polarized domains of the plasma membrane.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Hartzell C, Putzier I, Arreola J. Calcium-activated chloride channels. Annu Rev Physiol. 2005;67:719–758. - PubMed
-
- Yang YD, et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature. 2008;455:1210–1215. - PubMed
-
- Caputo A, et al. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science. 2008;322:590–594. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30NS055077/NS/NINDS NIH HHS/United States
- R56 NS042599/NS/NINDS NIH HHS/United States
- P30 EY006360/EY/NEI NIH HHS/United States
- NS42599/NS/NINDS NIH HHS/United States
- GM077569/GM/NIGMS NIH HHS/United States
- 5T32EY007092-25/EY/NEI NIH HHS/United States
- T32 EY007092/EY/NEI NIH HHS/United States
- R01 GM077569/GM/NIGMS NIH HHS/United States
- R01 GM060448/GM/NIGMS NIH HHS/United States
- P30 NS055077/NS/NINDS NIH HHS/United States
- R01 NS042599/NS/NINDS NIH HHS/United States
- P30EY006360/EY/NEI NIH HHS/United States
- R01 EY014852/EY/NEI NIH HHS/United States
- GM60448/GM/NIGMS NIH HHS/United States
- EY014852/EY/NEI NIH HHS/United States
- K12 GM000608/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
