

TNFR1-activated reactive oxidative species signals up-regulate osteogenic Msx2 programs in aortic myofibroblasts
- PMID: 22685265
- PMCID: PMC3404358
- DOI: 10.1210/en.2012-1216
TNFR1-activated reactive oxidative species signals up-regulate osteogenic Msx2 programs in aortic myofibroblasts
Abstract
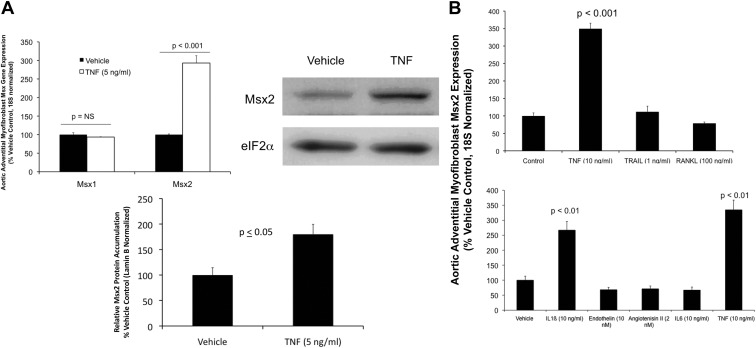
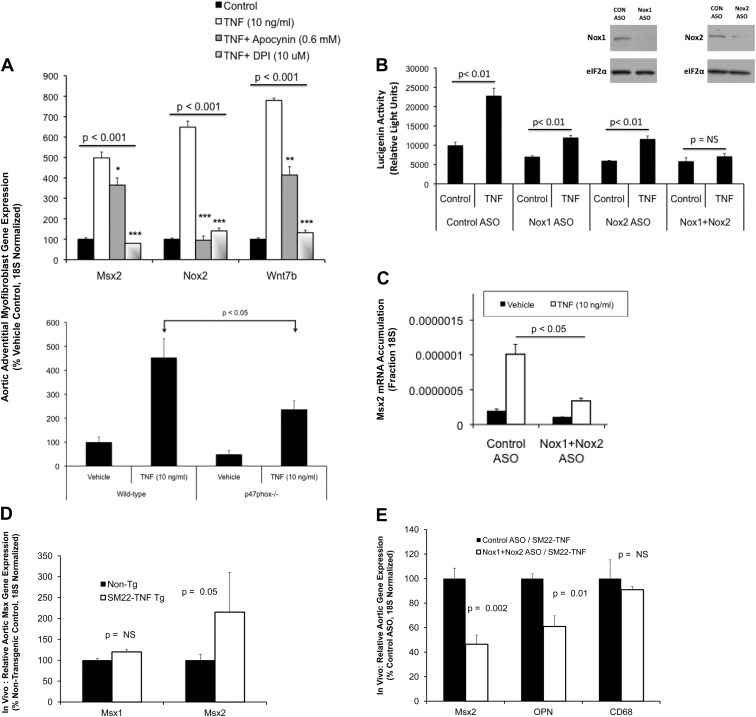
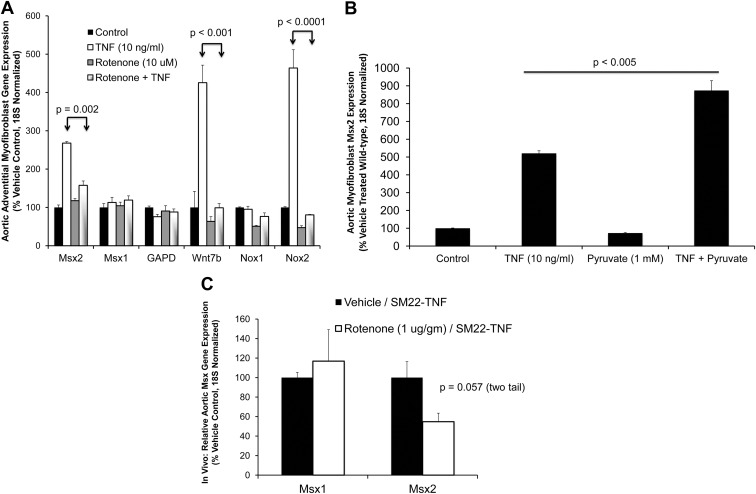
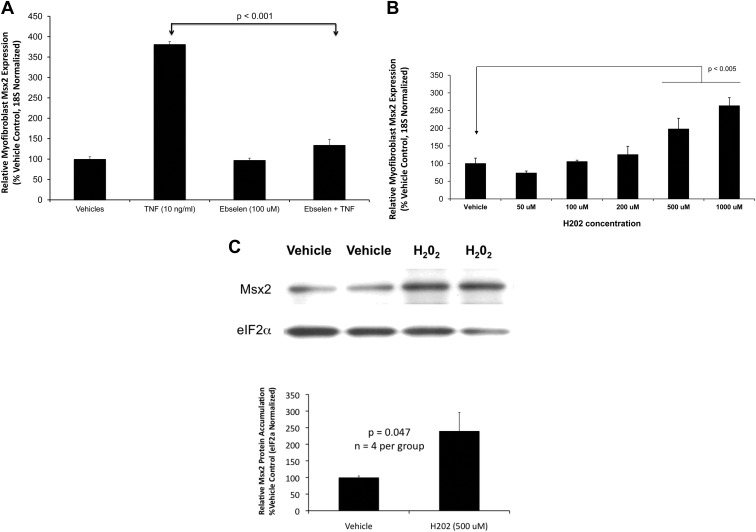
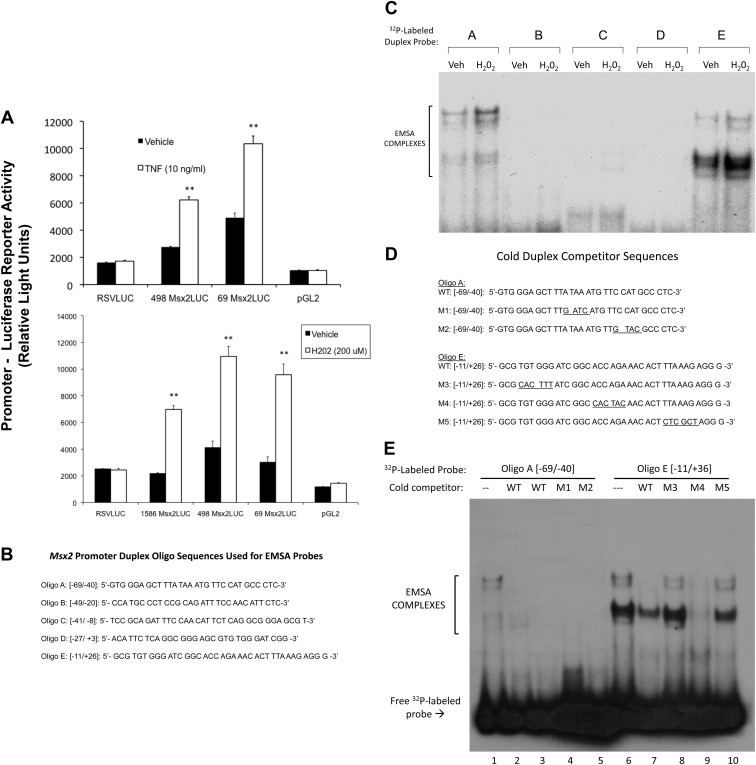
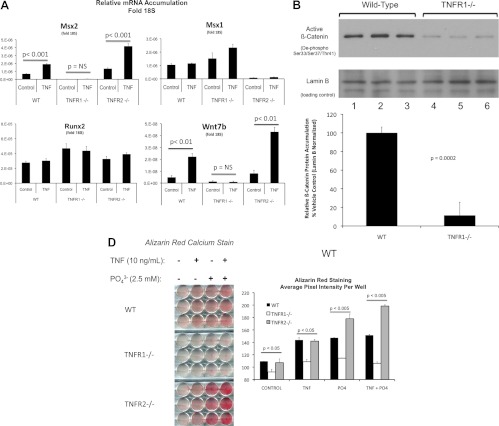
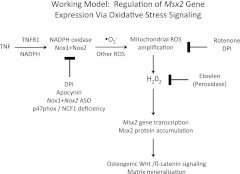
In LDLR(-/-) mice fed high-fat diabetogenic diets, osteogenic gene-regulatory programs are ectopically activated in vascular myofibroblasts and smooth muscle cells that promote arteriosclerotic calcium deposition. Msx2-Wnt signaling pathways previously identified as important for craniofacial skeletal development are induced in the vasculature by TNF, a prototypic cytokine mediator of the low-grade systemic inflammation of diabesity. To better understand this biology, we studied TNF actions on Msx2 in aortic myofibroblasts. TNF up-regulated Msx2 mRNA 4-fold within 3 h but did not regulate Msx1. Although IL-1β could also induce Msx2 expression, TNF-related apoptosis inducing ligand, receptor activator of nuclear factor-κB ligand, and IL-6 were inactive. Inhibition of nicotinamide adenine dinucleotide phosphate oxidase (Nox) activity and genetically induced Nox deficiency (p47phox(-/-)) reduced Msx2 induction, indicating contributions of reactive oxygen species (ROS) and redox signaling. Consistent with this, rotenone, an antagonist of mitochondrial complex I, inhibited TNF induction of Msx2 and Nox2, whereas pyruvate, an anapleurotic mitochondrial metabolic substrate, enhanced induction. Moreover, the glutathione peroxidase-mimetic ebselen abrogated this TNF response. Treatment of aortic myofibroblasts with hydrogen peroxide up-regulated Msx2 mRNA, promoter activity, and DNA-protein interactions. In vivo, SM22-TNF transgenic mice exhibit increased aortic Msx2 with no change in Msx1. Dosing SM22-TNF mice with either 20 ng/g Nox1 + 20 ng/g Nox2 antisense oligonucleotides or low-dose rotenone reduced arterial Msx2 expression. Aortic myofibroblasts from TNFR1(-/-) mice expressed levels of Msx2 that were 5% that of wild-type and were not inducible by TNF. Wnt7b and active β-catenin levels were also reduced. By contrast, TNF-inducible Msx2 expression was not reduced in TNFR2(-/-) cells. Finally, when cultured under mineralizing conditions, TNFR1(-/-) aortic myofibroblasts exhibited reduced calcification compared with wild-type and TNFR2(-/-) cells. Thus, ROS metabolism contributes to TNF induction of Msx2 and procalcific responses in myofibroblasts via TNFR1. Strategies that reduce vascular Nox- or mitochondrially activated ROS signals may prove useful in mitigating arteriosclerotic calcification.
Figures
References
-
- Greenwald SE. 2007. Ageing of the conduit arteries. J Pathol 211:157–172
-
- Safar ME, Boudier HS. 2005. Vascular development, pulse pressure, and the mechanisms of hypertension. Hypertension 46:205–209 - PubMed
-
- Towler DA, Bidder M, Latifi T, Coleman T, Semenkovich CF. 1998. Diet-induced diabetes activates an osteogenic gene regulatory program in the aortas of low density lipoprotein receptor-deficient mice. J Biol Chem 273:30427–30434 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
