

Elongation factor 1A is the target of growth inhibition in yeast caused by Legionella pneumophila glucosyltransferase Lgt1
- PMID: 22685293
- PMCID: PMC3406686
- DOI: 10.1074/jbc.M112.372672
Elongation factor 1A is the target of growth inhibition in yeast caused by Legionella pneumophila glucosyltransferase Lgt1
Abstract
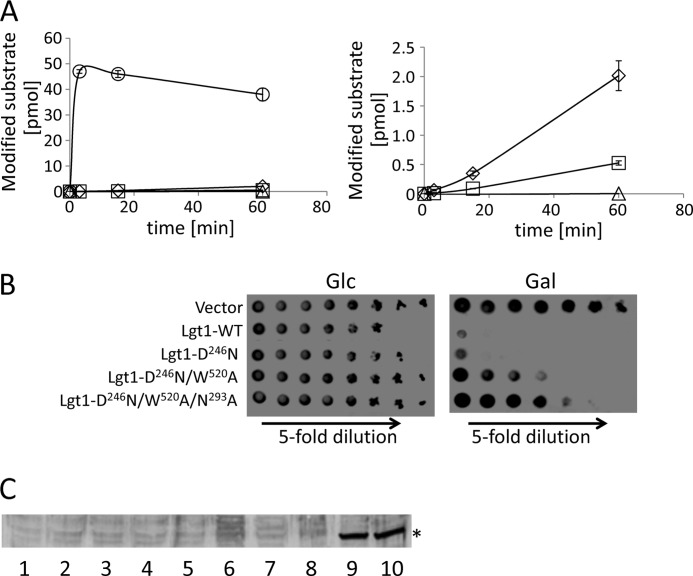
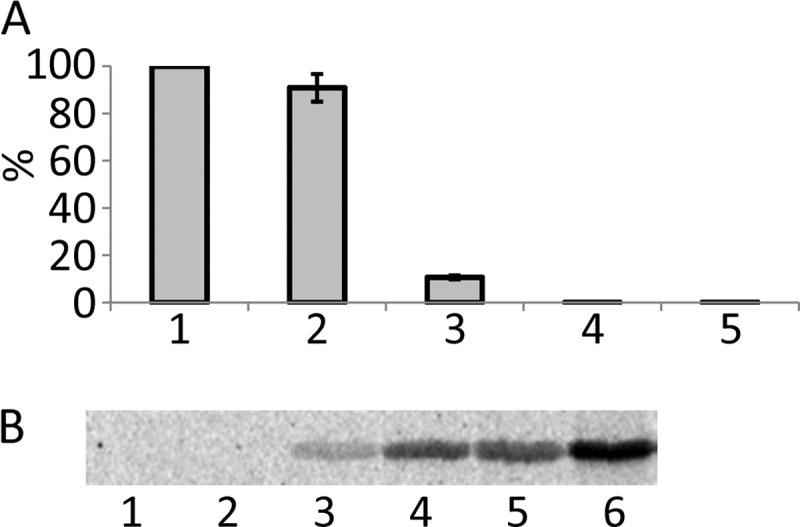
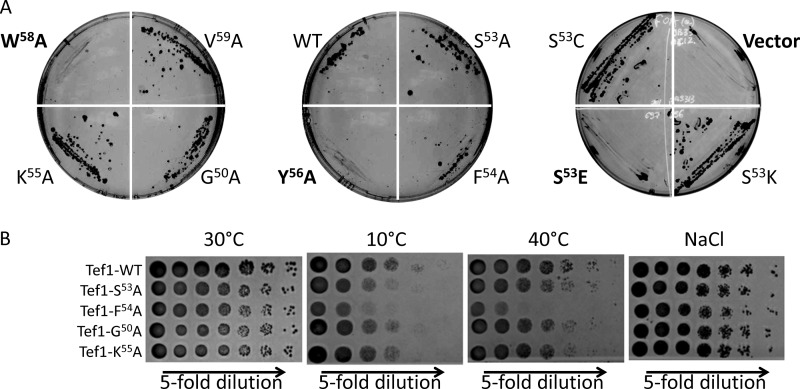
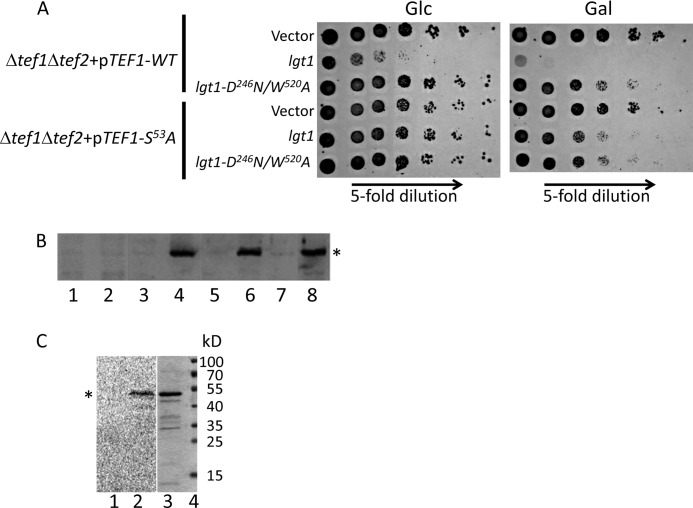
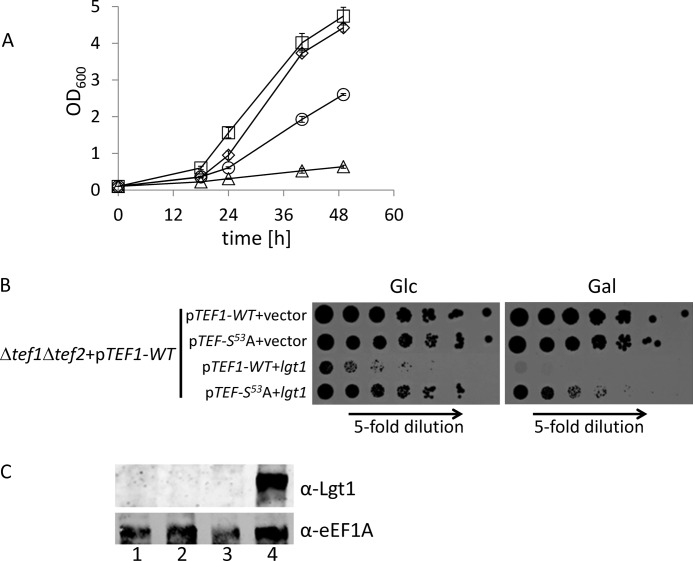
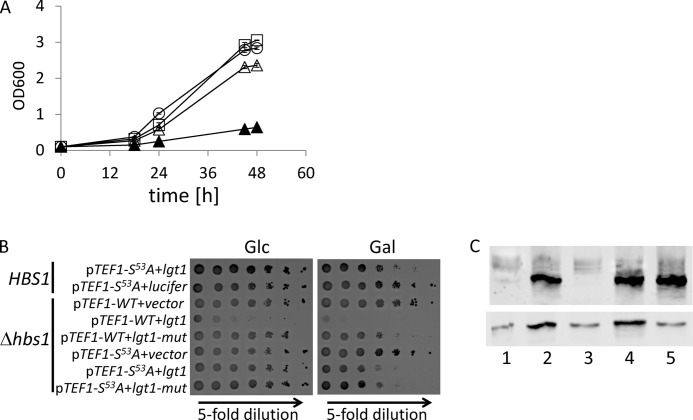
Legionella is a pathogenic Gram-negative bacterium that can multiply inside of eukaryotic cells. It translocates numerous bacterial effector proteins into target cells to transform host phagocytes into a niche for replication. One effector of Legionella pneumophila is the glucosyltransferase Lgt1, which modifies serine 53 in mammalian elongation factor 1A (eEF1A), resulting in inhibition of protein synthesis and cell death. Here, we demonstrate that similar to mammalian cells, Lgt1 was severely toxic when produced in yeast and effectively inhibited in vitro protein synthesis. Saccharomyces cerevisiae strains, which were deleted of endogenous eEF1A but harbored a mutant eEF1A not glucosylated by Lgt1, were resistant toward the bacterial effector. In contrast, deletion of Hbs1, which is also an in vitro substrate of the glucosyltransferase, did not influence the toxic effects of Lgt1. Serial mutagenesis in yeast showed that Phe(54), Tyr(56) and Trp(58), located immediately downstream of serine 53 of eEF1A, are essential for the function of the elongation factor. Replacement of serine 53 by glutamic acid, mimicking phosphorylation, produced a non-functional eEF1A, which failed to support growth of S. cerevisiae. Our data indicate that Lgt1-induced lethal effect in yeast depends solely on eEF1A. The region of eEF1A encompassing serine 53 plays a critical role in functioning of the elongation factor.
Figures

References
-
- McDade J. E., Shepard C. C., Fraser D. W., Tsai T. R., Redus M. A., Dowdle W. R. (1977) Legionnaires' disease: Isolation of a bacterium and demonstration of its role in other respiratory disease. N. Engl. J. Med. 297, 1197–1203 - PubMed
-
- Fraser D. W., Tsai T. R., Orenstein W., Parkin W. E., Beecham H. J., Sharrar R. G., Harris J., Mallison G. F., Martin S. M., McDade J. E., Shepard C. C., Brachman P. S. (1977) Legionnaires' disease: Description of an epidemic of pneumonia. N. Engl. J. Med. 297, 1189–1197 - PubMed
-
- Jules M., Buchrieser C. (2007) Legionella pneumophila adaptation to intracellular life and the host response: Clues from genomics and transcriptomics. FEBS Lett. 581, 2829–2838 - PubMed
-
- Hubber A., Roy C. R. (2010) Modulation of host cell function by Legionella pneumophila type IV effectors. Annu. Rev. Cell Dev. Biol. 26, 261–283 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
