

Biological significance of nuclear localization of mitogen-activated protein kinase Pmk1 in fission yeast
- PMID: 22685296
- PMCID: PMC3406687
- DOI: 10.1074/jbc.M112.345611
Biological significance of nuclear localization of mitogen-activated protein kinase Pmk1 in fission yeast
Abstract
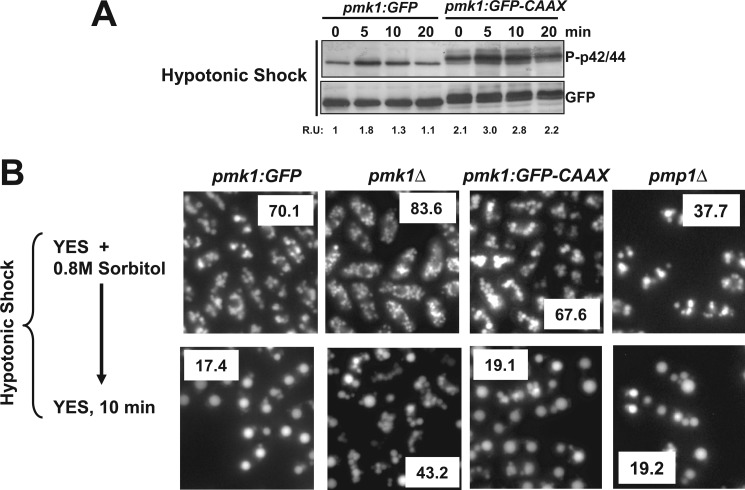
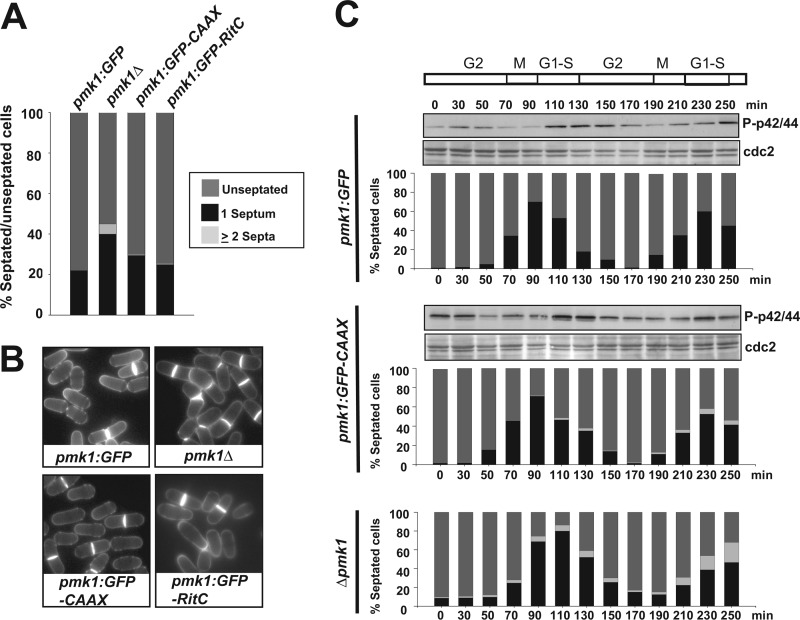
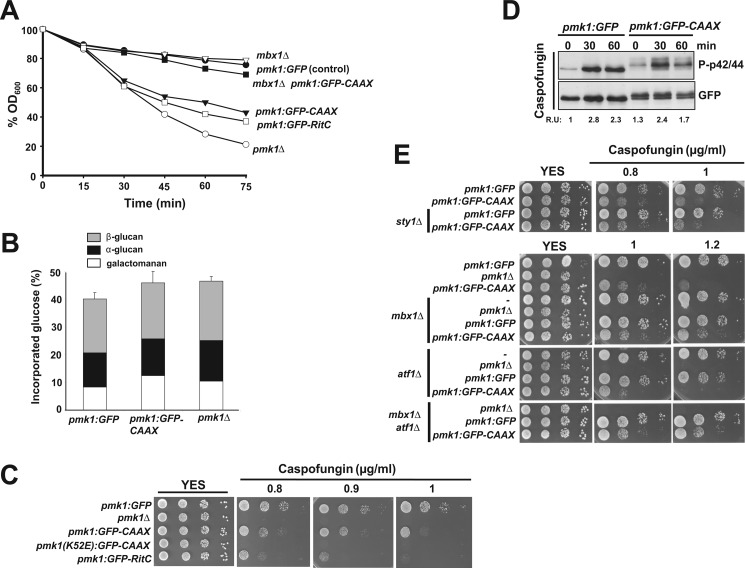
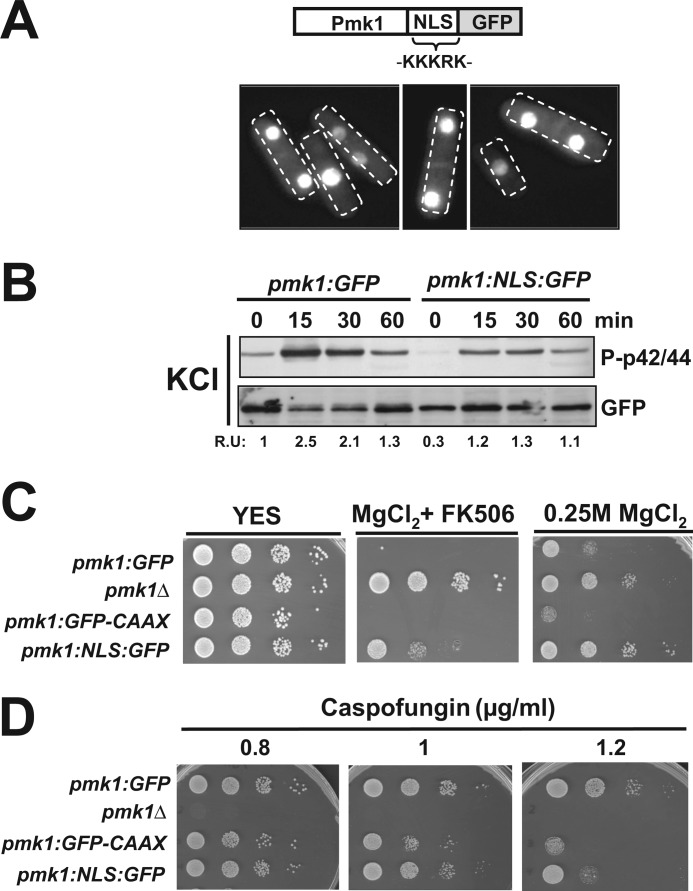
Mitogen-activated protein kinase (MAPK) signaling pathways play a fundamental role in the response of eukaryotic cells to environmental changes. Also, much evidence shows that the stimulus-dependent nuclear targeting of this class of regulatory kinases is crucial for adequate regulation of distinct cellular events. In the fission yeast Schizosaccharomyces pombe, the cell integrity MAPK pathway, whose central element is the MAPK Pmk1, regulates multiple processes such as cell wall integrity, vacuole fusion, cytokinesis, and ionic homeostasis. In non-stressed cells Pmk1 is constitutively localized in both cytoplasm and nucleus, and its localization pattern appears unaffected by its activation status or in response to stress, thus questioning the biological significance of the presence of this MAPK into the nucleus. We have addressed this issue by characterizing mutants expressing Pmk1 versions excluded from the cell nucleus and anchored to the plasma membrane in different genetic backgrounds. Although nuclear Pmk1 partially regulates cell wall integrity at a transcriptional level, membrane-tethered Pmk1 performs many of the biological functions assigned to wild type MAPK like regulation of chloride homeostasis, vacuole fusion, and cellular separation. However, we found that down-regulation of nuclear Pmk1 by MAPK phosphatases induced by the stress activated protein kinase pathway is important for the fine modulation of extranuclear Pmk1 activity. These results highlight the importance of the control of MAPK activity at subcellular level.
Figures
Similar articles
-
Stress-induced response, localization, and regulation of the Pmk1 cell integrity pathway in Schizosaccharomyces pombe.J Biol Chem. 2006 Jan 27;281(4):2033-43. doi: 10.1074/jbc.M506467200. Epub 2005 Nov 16. J Biol Chem. 2006. PMID: 16291757
-
Role of the fission yeast cell integrity MAPK pathway in response to glucose limitation.BMC Microbiol. 2013 Feb 11;13:34. doi: 10.1186/1471-2180-13-34. BMC Microbiol. 2013. PMID: 23398982 Free PMC article.
-
Geranylgeranyltransferase Cwg2-Rho4/Rho5 module is implicated in the Pmk1 MAP kinase-mediated cell wall integrity pathway in fission yeast.Genes Cells. 2015 Apr;20(4):310-23. doi: 10.1111/gtc.12222. Epub 2015 Feb 5. Genes Cells. 2015. PMID: 25651781
-
[Cell surface protein Ecm33 is involved in negative feedback regulation of MAP kinase signalling and development of the in vivo real-time monitoring of MAP kinase signalling].Yakugaku Zasshi. 2011;131(8):1195-200. doi: 10.1248/yakushi.131.1195. Yakugaku Zasshi. 2011. PMID: 21804323 Review. Japanese.
-
[The Molecular Basis of Drug Discovery Targeting the Regulatory Mechanism of MAPK Signaling via the Spatial Regulation of RNA-binding Proteins].Yakugaku Zasshi. 2019;139(1):7-12. doi: 10.1248/yakushi.18-00189. Yakugaku Zasshi. 2019. PMID: 30606933 Review. Japanese.
Cited by
-
Rho2 palmitoylation is required for plasma membrane localization and proper signaling to the fission yeast cell integrity mitogen- activated protein kinase pathway.Mol Cell Biol. 2014 Jul;34(14):2745-59. doi: 10.1128/MCB.01515-13. Mol Cell Biol. 2014. PMID: 24820419 Free PMC article.
-
A novel checkpoint pathway controls actomyosin ring constriction trigger in fission yeast.Elife. 2020 Oct 26;9:e59333. doi: 10.7554/eLife.59333. Elife. 2020. PMID: 33103994 Free PMC article.
-
Pck2 association with the plasma membrane and efficient response of the cell integrity pathway require regulation of PI4P homeostasis by exomer.Open Biol. 2024 Nov;14(11):240101. doi: 10.1098/rsob.240101. Epub 2024 Nov 13. Open Biol. 2024. PMID: 39540318 Free PMC article.
-
Cotton GhMKK1 induces the tolerance of salt and drought stress, and mediates defence responses to pathogen infection in transgenic Nicotiana benthamiana.PLoS One. 2013 Jul 3;8(7):e68503. doi: 10.1371/journal.pone.0068503. Print 2013. PLoS One. 2013. PMID: 23844212 Free PMC article.
-
Fission yeast cell wall biosynthesis and cell integrity signalling.Cell Surf. 2018 Oct 17;4:1-9. doi: 10.1016/j.tcsw.2018.10.001. eCollection 2018 Dec. Cell Surf. 2018. PMID: 32743131 Free PMC article. Review.
References
-
- Kim E. K., Choi E. J. (2010) Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 1802, 396–405 - PubMed
-
- Qi M., Elion E. A. (2005) MAP kinase pathways. J. Cell Sci. 118, 3569–3572 - PubMed
-
- Krisak L., Strich R., Winters R. S., Hall J. P., Mallory M. J., Kreitzer D., Tuan R. S., Winter E. (1994) SMK1, a developmentally regulated MAP kinase, is required for spore wall assembly in Saccharomyces cerevisiae. Genes Dev. 8, 2151–2161 - PubMed
-
- Shiozaki K., Russell P. (1995) Cell cycle control linked to extracellular environment by MAP kinase pathway in fission yeast. Nature 378, 739–743 - PubMed
-
- Toda T., Dhut S., Superti-Furga G., Gotoh Y., Nishida E., Sugiura R., Kuno T. (1996) The fission yeast pmk1+ gene encodes a novel mitogen-activated protein kinase homolog that regulates cell integrity and functions coordinately with the protein kinase C pathway. Mol. Cell. Biol. 16, 6752–6764 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
