

Anopheles Imd pathway factors and effectors in infection intensity-dependent anti-Plasmodium action
- PMID: 22685401
- PMCID: PMC3369948
- DOI: 10.1371/journal.ppat.1002737
Anopheles Imd pathway factors and effectors in infection intensity-dependent anti-Plasmodium action
Abstract
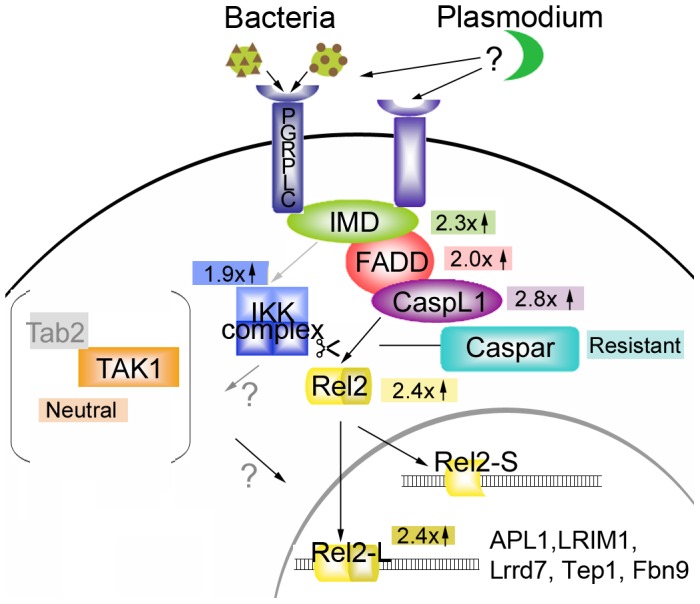
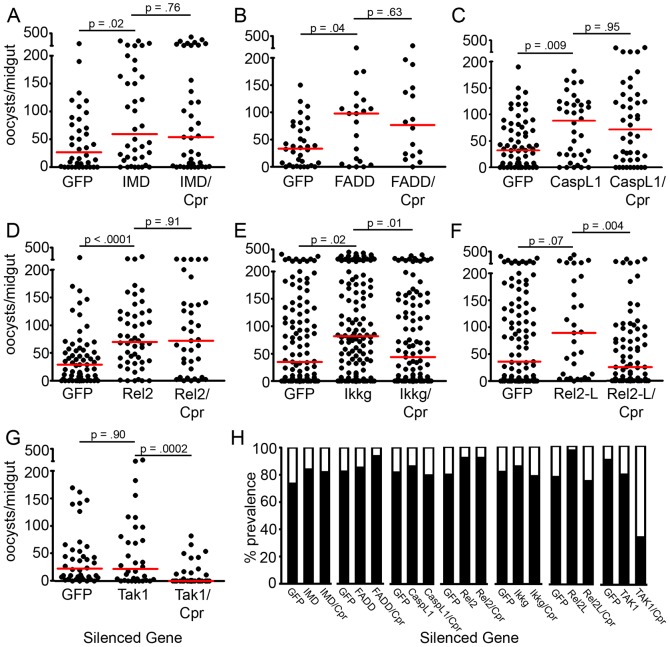
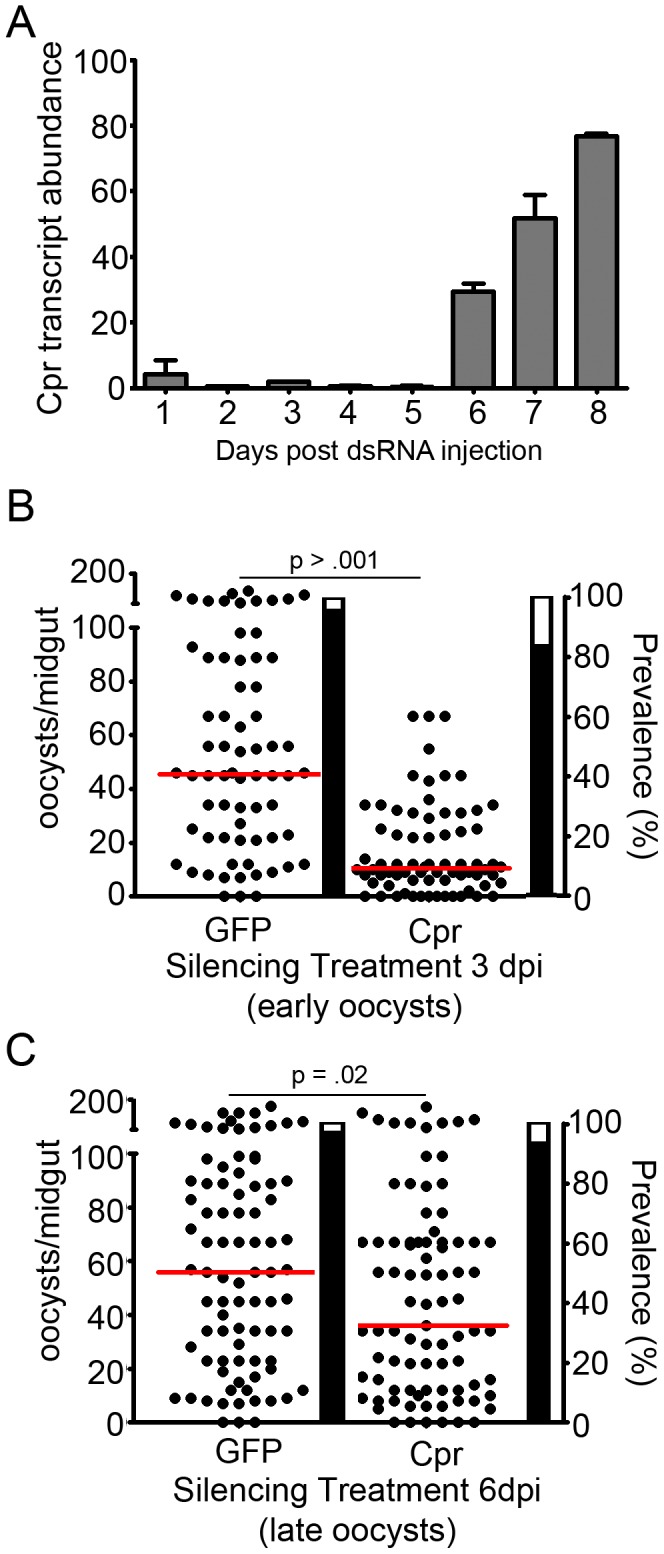
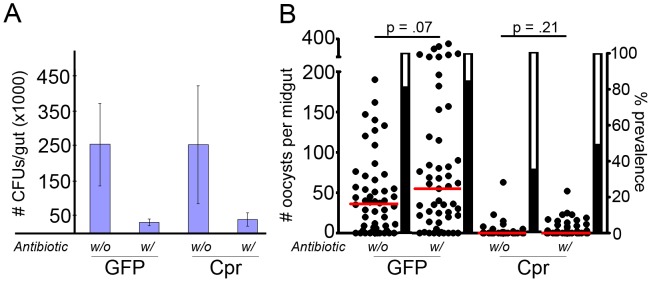
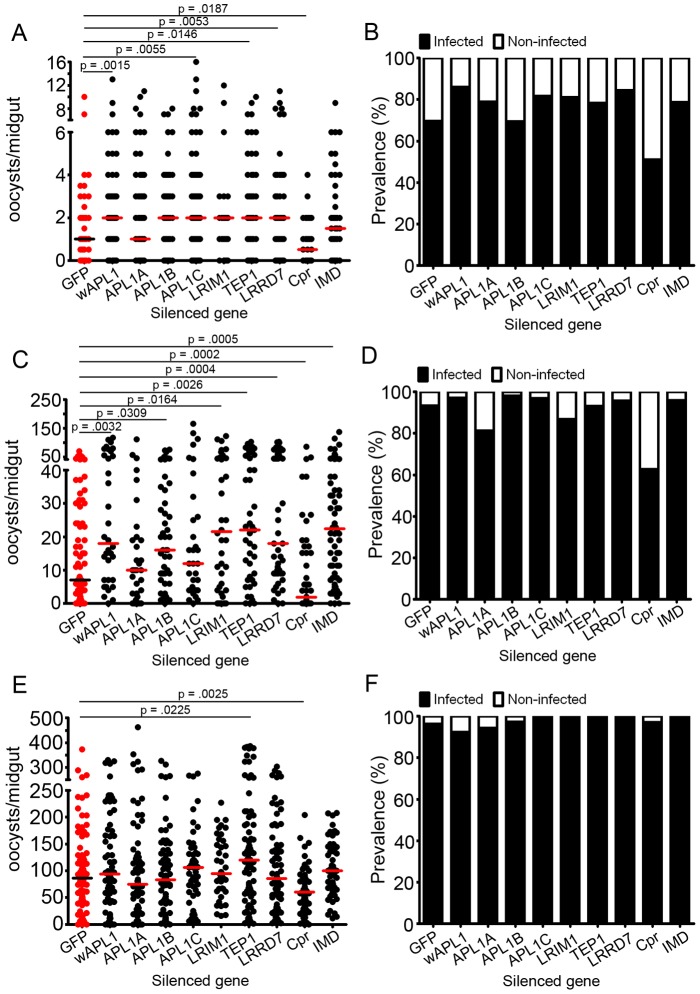
The Anopheles gambiae immune response against Plasmodium falciparum, an etiological agent of human malaria, has been identified as a source of potential anti-Plasmodium genes and mechanisms to be exploited in efforts to control the malaria transmission cycle. One such mechanism is the Imd pathway, a conserved immune signaling pathway that has potent anti-P. falciparum activity. Silencing the expression of caspar, a negative regulator of the Imd pathway, or over-expressing rel2, an Imd pathway-controlled NFkappaB transcription factor, confers a resistant phenotype on A. gambiae mosquitoes that involves an array of immune effector genes. However, unexplored features of this powerful mechanism that may be essential for the implementation of a malaria control strategy still remain. Using RNA interference to singly or dually silence caspar and other components of the Imd pathway, we have identified genes participating in the anti-Plasmodium signaling module regulated by Caspar, each of which represents a potential target to achieve over-activation of the pathway. We also determined that the Imd pathway is most potent against the parasite's ookinete stage, yet also has reasonable activity against early oocysts and lesser activity against late oocysts. We further demonstrated that caspar silencing alone is sufficient to induce a robust anti-P. falciparum response even in the relative absence of resident gut microbiota. Finally, we established the relevance of the Imd pathway components and regulated effectors TEP1, APL1, and LRIM1 in parasite infection intensity-dependent defense, thereby shedding light on the relevance of laboratory versus natural infection intensity models. Our results highlight the physiological considerations that are integral to a thoughtful implementation of Imd pathway manipulation in A. gambiae as part of an effort to limit the malaria transmission cycle, and they reveal a variety of previously unrecognized nuances in the Imd-directed immune response against P. falciparum.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- World Health Organization. Global Malaria Control and Elimination: report of a technical review. Geneva, Switzerland. 2008.
-
- World Health Organization. World Malaria Report. Geneva, Switzerland. 2010.
-
- Frolet C, Thoma M, Blandin S, Hoffmann JA, Levashina EA. Boosting NF-κB-dependent basal immunity of Anopheles gambiae aborts development of Plasmodium berghei. Immunity. 2006;25:685. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
