

Specific evolution of F1-like ATPases in mycoplasmas
- PMID: 22685606
- PMCID: PMC3369863
- DOI: 10.1371/journal.pone.0038793
Specific evolution of F1-like ATPases in mycoplasmas
Abstract
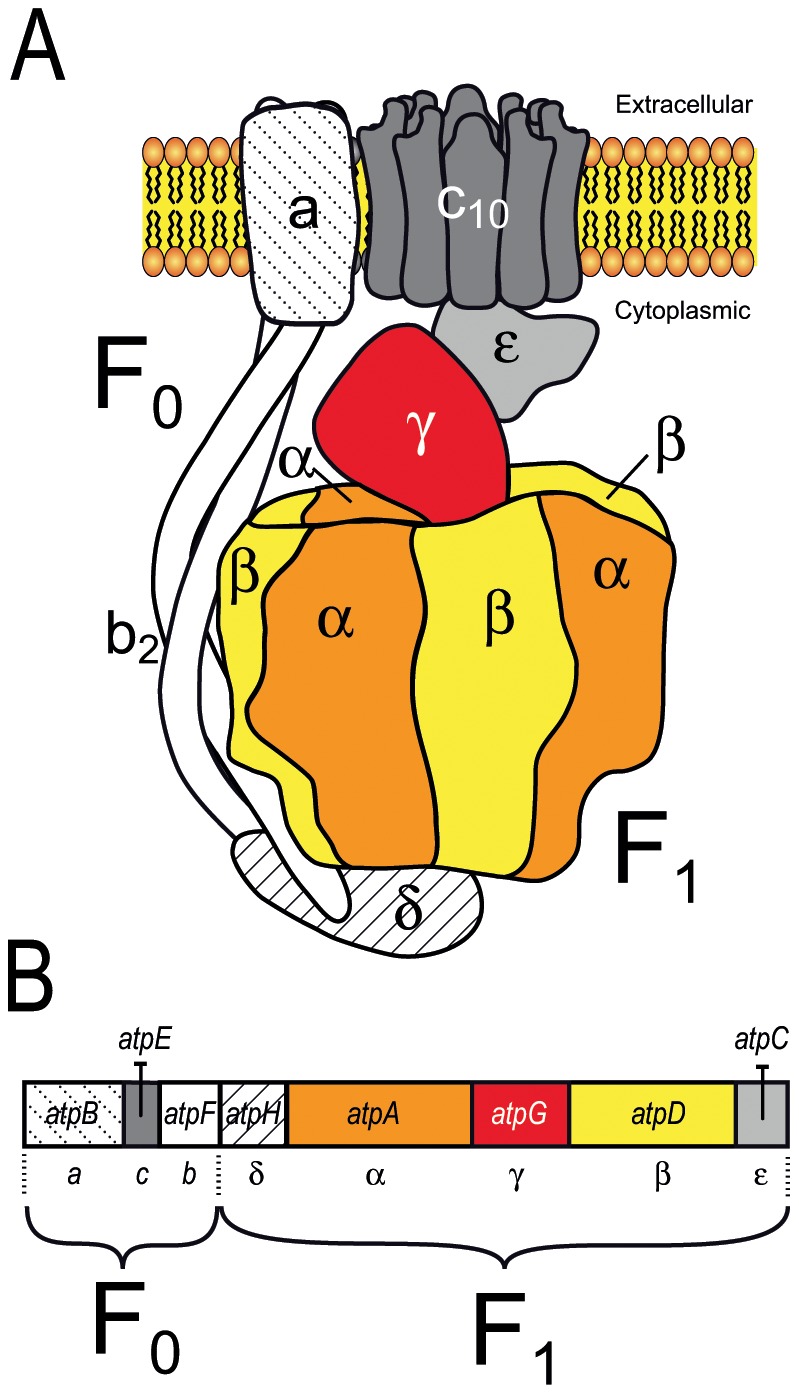
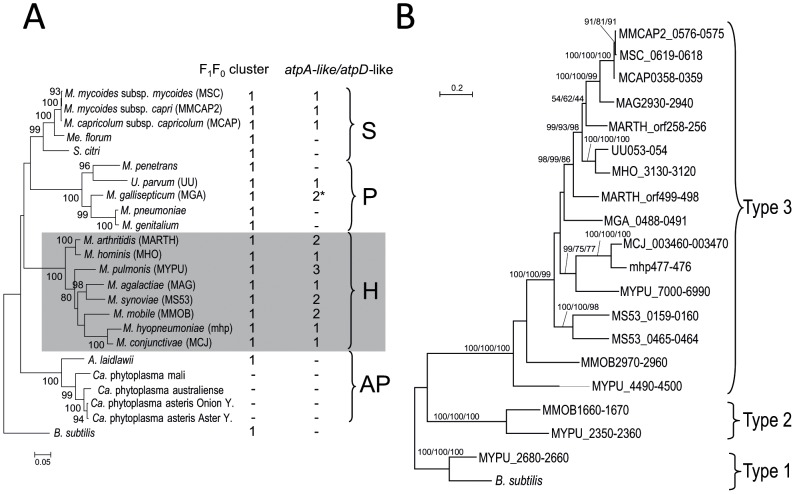
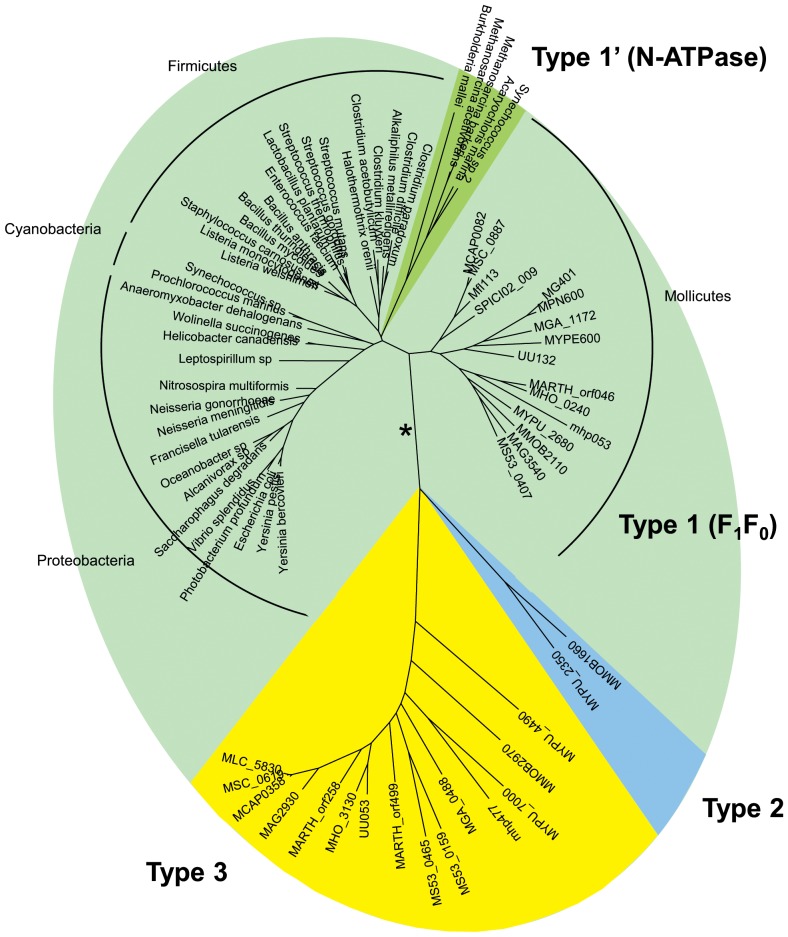
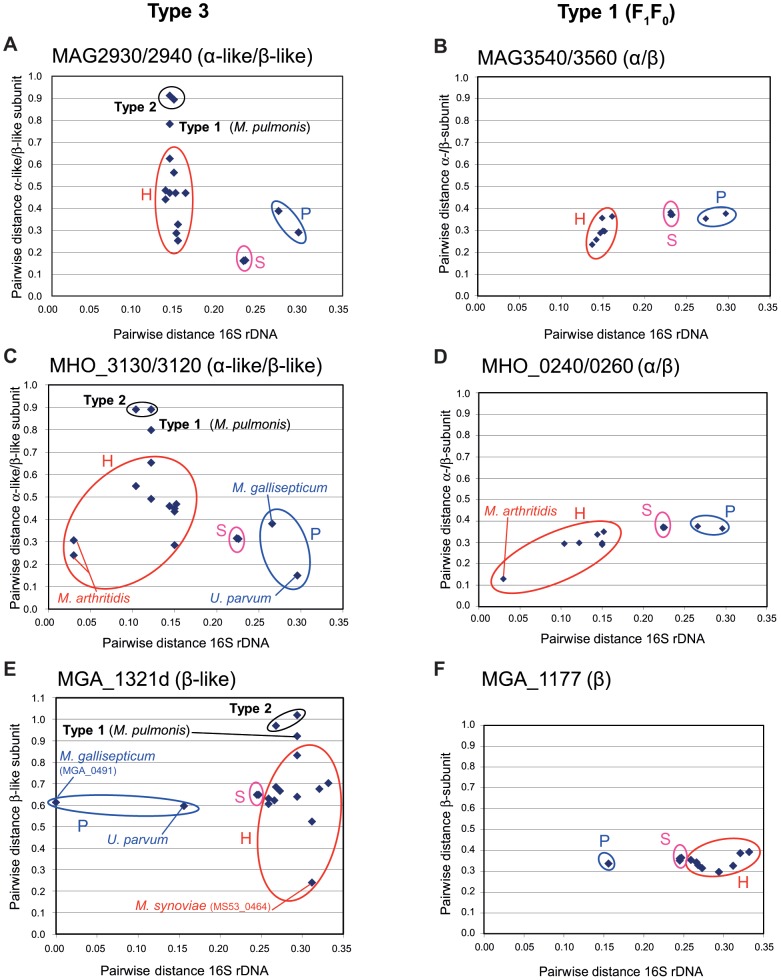
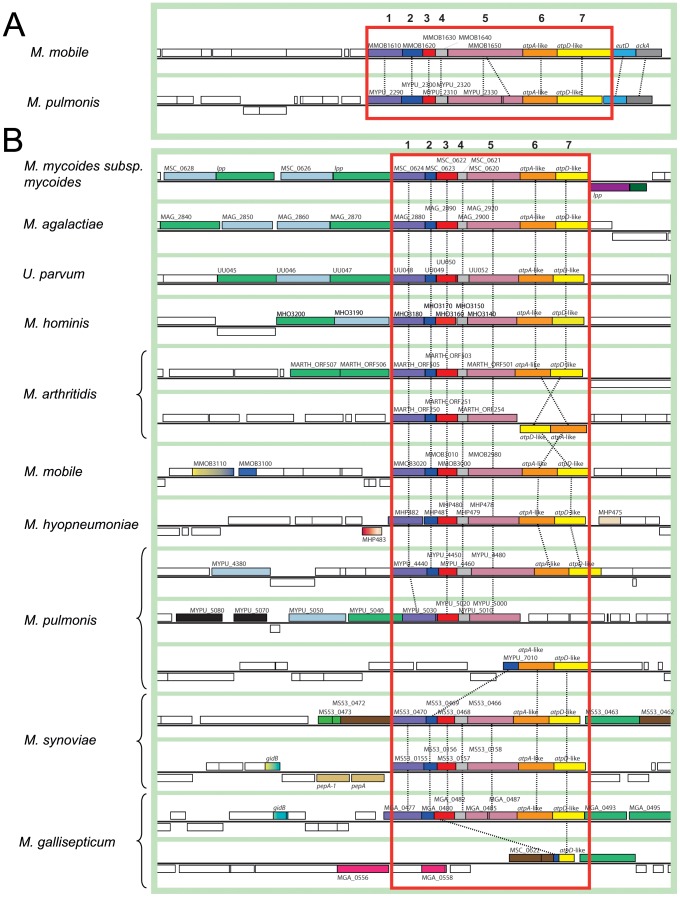
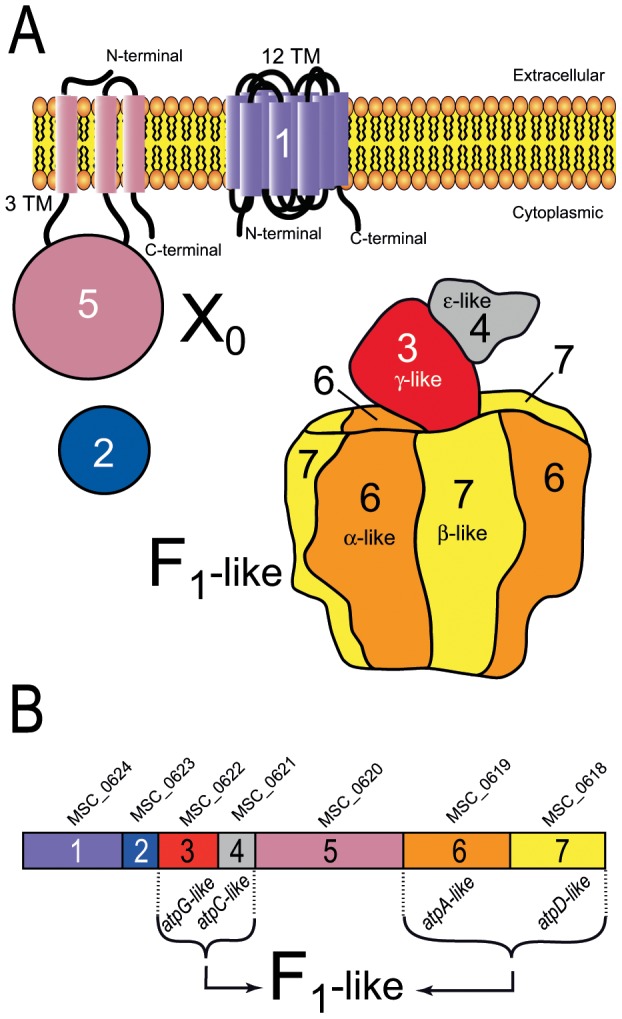
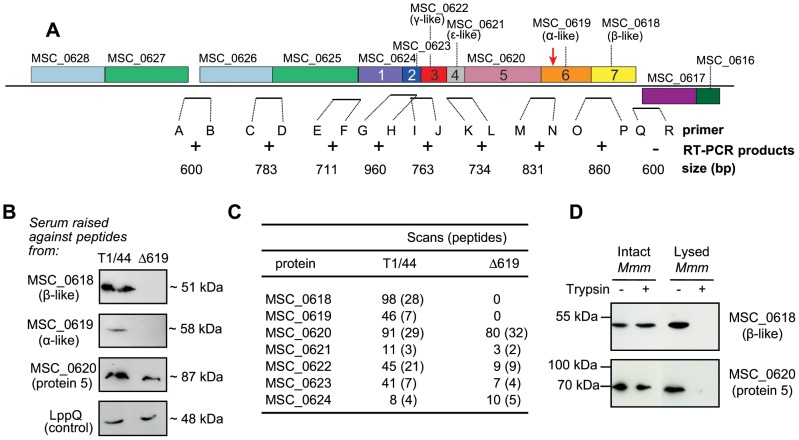
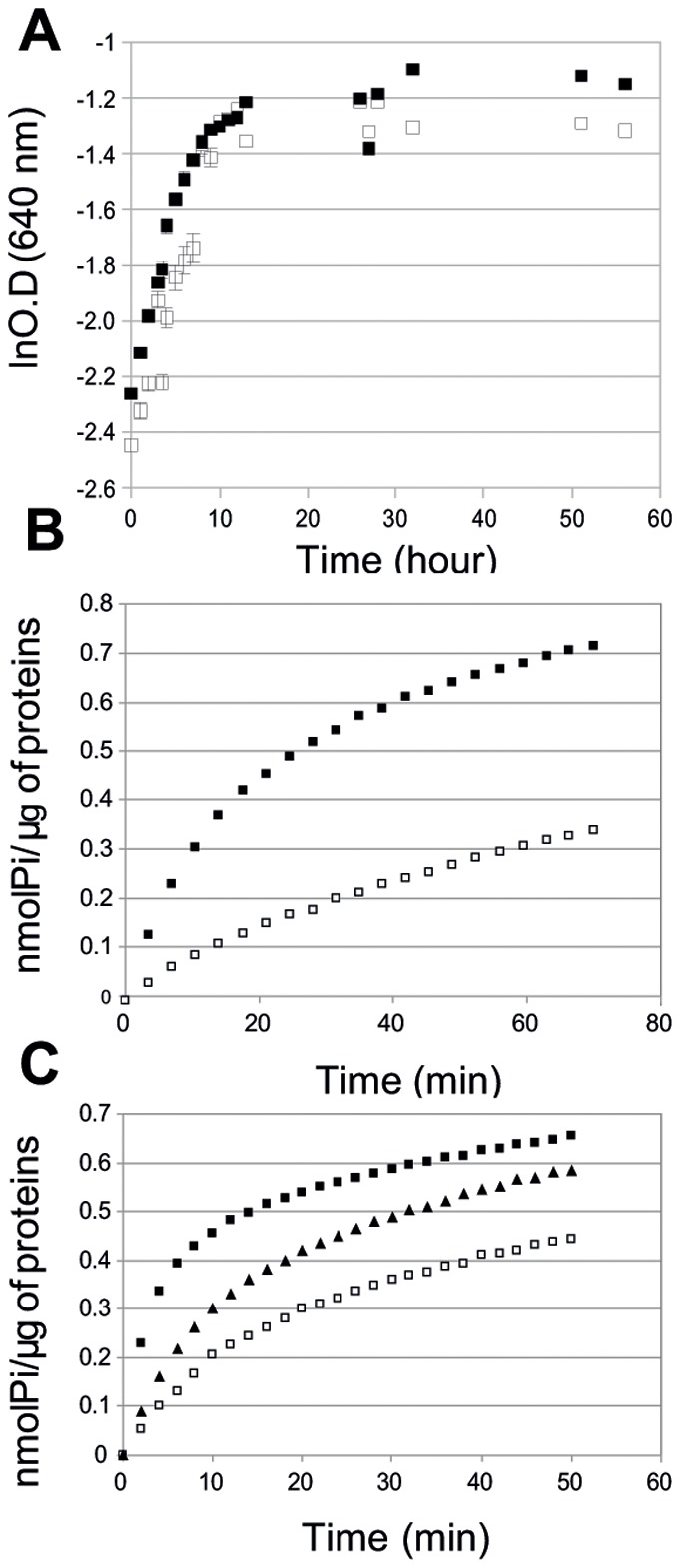
F(1)F(0) ATPases have been identified in most bacteria, including mycoplasmas which have very small genomes associated with a host-dependent lifestyle. In addition to the typical operon of eight genes encoding genuine F(1)F(0) ATPase (Type 1), we identified related clusters of seven genes in many mycoplasma species. Four of the encoded proteins have predicted structures similar to the α, β, γ and ε subunits of F(1) ATPases and could form an F(1)-like ATPase. The other three proteins display no similarity to any other known proteins. Two of these proteins are probably located in the membrane, as they have three and twelve predicted transmembrane helices. Phylogenomic studies identified two types of F(1)-like ATPase clusters, Type 2 and Type 3, characterized by a rapid evolution of sequences with the conservation of structural features. Clusters encoding Type 2 and Type 3 ATPases were assumed to originate from the Hominis group of mycoplasmas. We suggest that Type 3 ATPase clusters may spread to other phylogenetic groups by horizontal gene transfer between mycoplasmas in the same host, based on phylogeny and genomic context. Functional analyses in the ruminant pathogen Mycoplasma mycoides subsp. mycoides showed that the Type 3 cluster genes were organized into an operon. Proteomic analyses demonstrated that the seven encoded proteins were produced during growth in axenic media. Mutagenesis and complementation studies demonstrated an association of the Type 3 cluster with a major ATPase activity of membrane fractions. Thus, despite their tendency toward genome reduction, mycoplasmas have evolved and exchanged specific F(1)-like ATPases with no known equivalent in other bacteria. We propose a model, in which the F(1)-like structure is associated with a hypothetical X(0) sector located in the membrane of mycoplasma cells.
Conflict of interest statement
Figures
References
-
- Sirand-Pugnet P, Citti C, Barre A, Blanchard A. Evolution of mollicutes: down a bumpy road with twists and turns. Res Microbiol. 2007;158:766. - PubMed
-
- Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B. Toward automatic reconstruction of a highly resolved tree of life. Science. 2006;311:1287. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
