

Review
doi: 10.1039/c2np20039b.
Epub 2012 Jun 12.
The structural biology of enzymes involved in natural product glycosylation
Affiliations
- PMID: 22688446
- PMCID: PMC3627186
- DOI: 10.1039/c2np20039b
Item in Clipboard
Review
The structural biology of enzymes involved in natural product glycosylation
Nat Prod Rep.
2012 Oct.
Abstract
The glycosylation of microbial natural products often dramatically influences the biological and/or pharmacological activities of the parental metabolite. Over the past decade, crystal structures of several enzymes involved in the biosynthesis and attachment of novel sugars found appended to natural products have emerged. In many cases, these studies have paved the way to a better understanding of the corresponding enzyme mechanism of action and have served as a starting point for engineering variant enzymes to facilitate to production of differentially-glycosylated natural products. This review specifically summarizes the structural studies of bacterial enzymes involved in biosynthesis of novel sugar nucleotides.
Figures
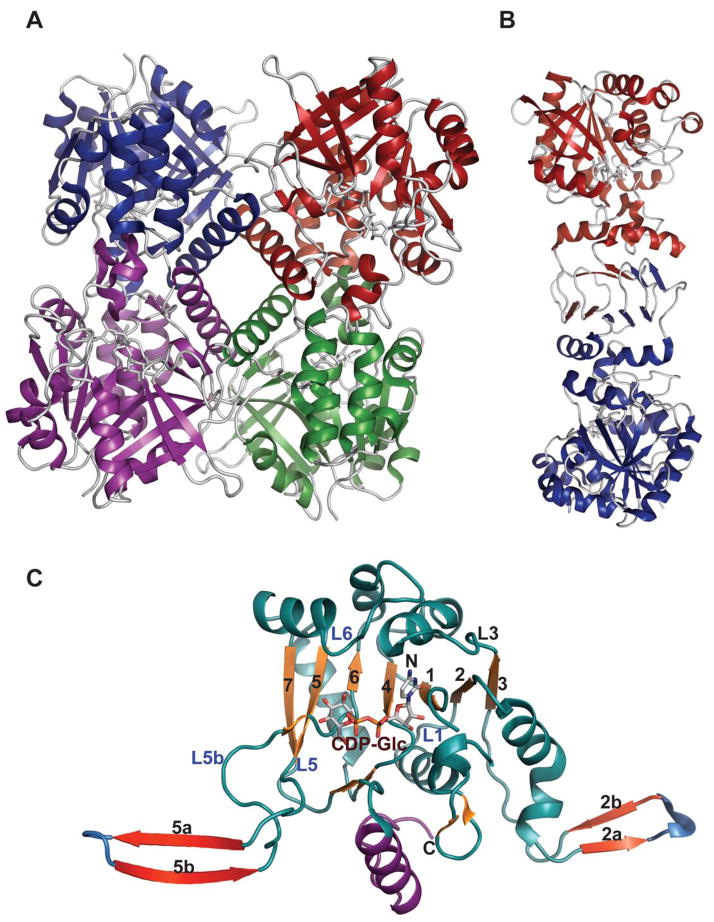
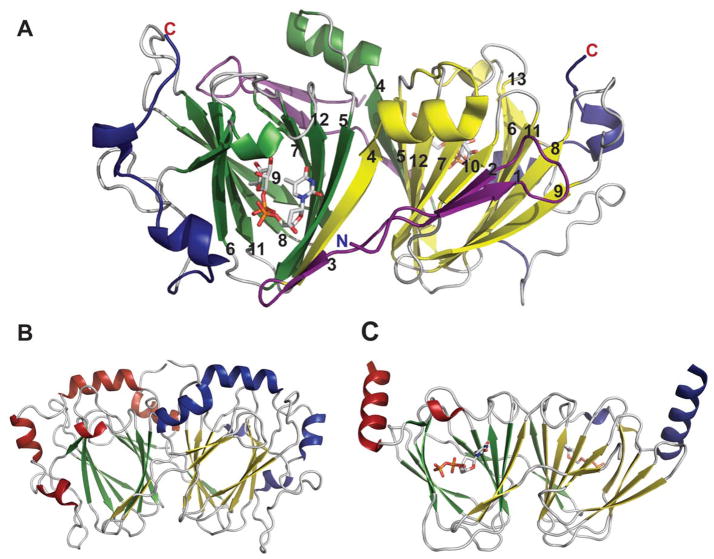
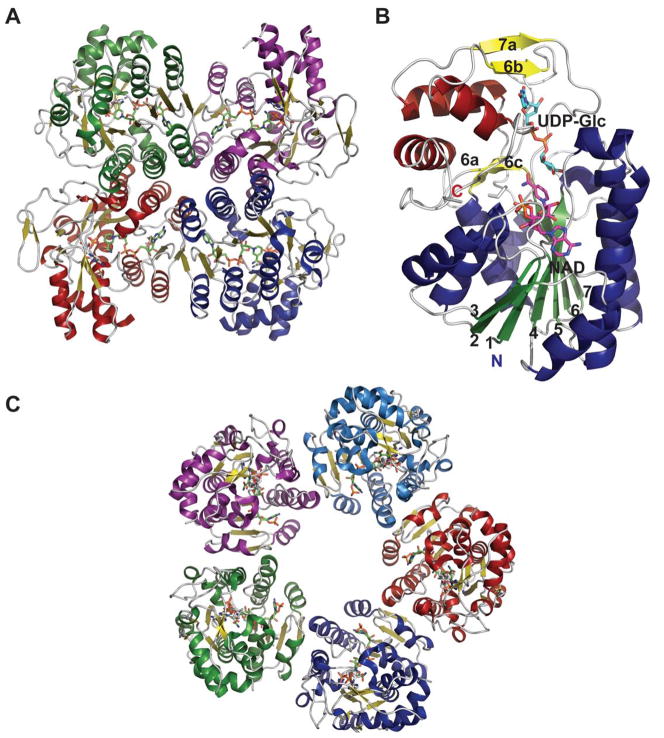
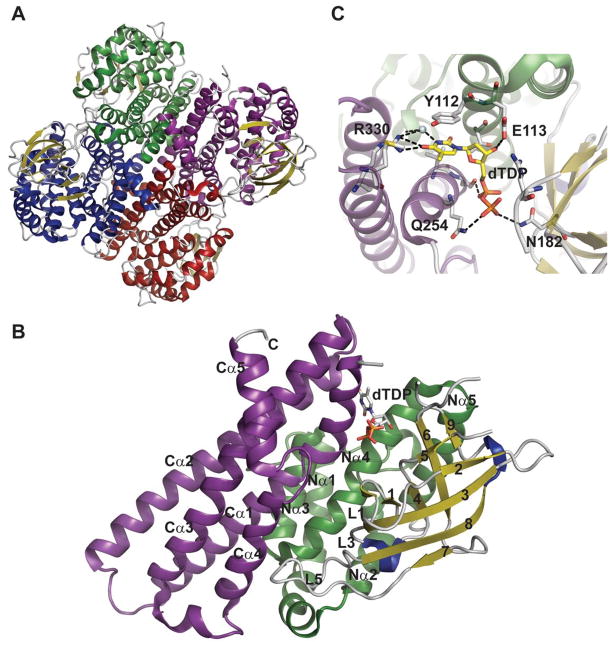
Representative global structural features of nucleotidyltransferases A. Tetramer of dTDP-glucose pyrophosphorylase (RmlA) from Salmonella enterica where each monomer is represented by a distinct color (PDB 1IIN). B. Dimer of GDP-mannose pyrophosphorylase (GMPase) from Thermotoga maritime where each monomer is represented by a distinct color. (PDB 2X5Z). C. Monomer of CDP-glucose pyrophosphorylase (CGPase) from Salmonella typhi (PDB 1TZF) with secondary structures of core β-sheet and variable regions numbered. The core N-terminal Rossmann fold is colored in green and orange (β-strands), the variable regions are colored blue and red, CDP-Glucose is colored grey, the C-terminal domain is purple and the letters ‘C’ and ‘N’ represent the C- and N-terminus of the enzyme, respectively.
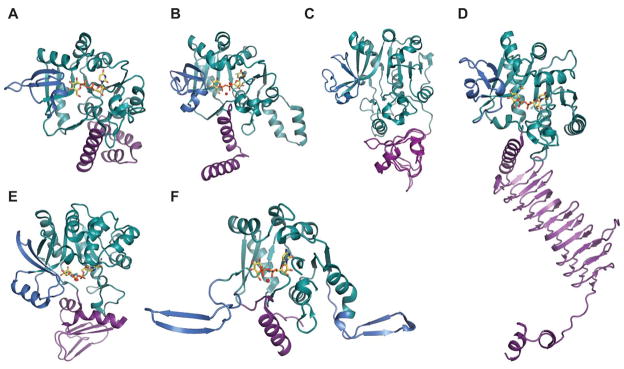
Representative monomeric features of nucleotidyltransferases. A. dTDP-glucose pyrophosphorylase RmlA from Salmonella enterica (PDB 1IIN). B. UDP-glucose pyrophosphorylase UGPase from Escherichia coli (PDB 2E3D). C. ADP-glucose pyrophosphorylase AGPase from Agrobacterium tumefaciens (PDB 3BRK). D.
N-acetylglucosamine diphosphorylase uridylyltransferase GlmU from Mycobacterium tuberculosis.
E. GDP-mannose pyrophosphorylase GMPase from Thermotoga maritime (PDB 2X5Z). F. CDP-glucose pyrophosphorylase CGPase from Salmonella typhi (PDB 1TZF). Rossmann fold is represented in green, variable regions colored blue, C-terminal domain in purple, ligands in yellow and Mg2+ as red sphere.
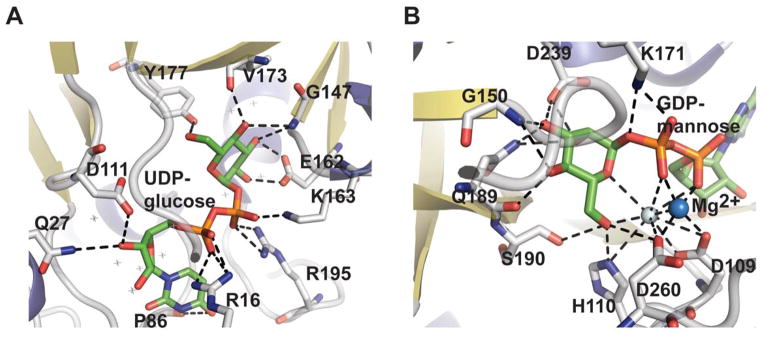
Nucleotidyltransferase active-site interactions. A. dTDP-glucose pyrophosphorylase RmlA from Salmonella enterica bound to UDP-glucose (PDB 1IIN). B. GDP-mannose pyrophosphorylase GMPase from Thermotoga maritime bound to GDP-mannose (PDB 2X5Z). NDP-sugar is colored green, Mg2+ is represented by a blue sphere, and water by a grey sphere.
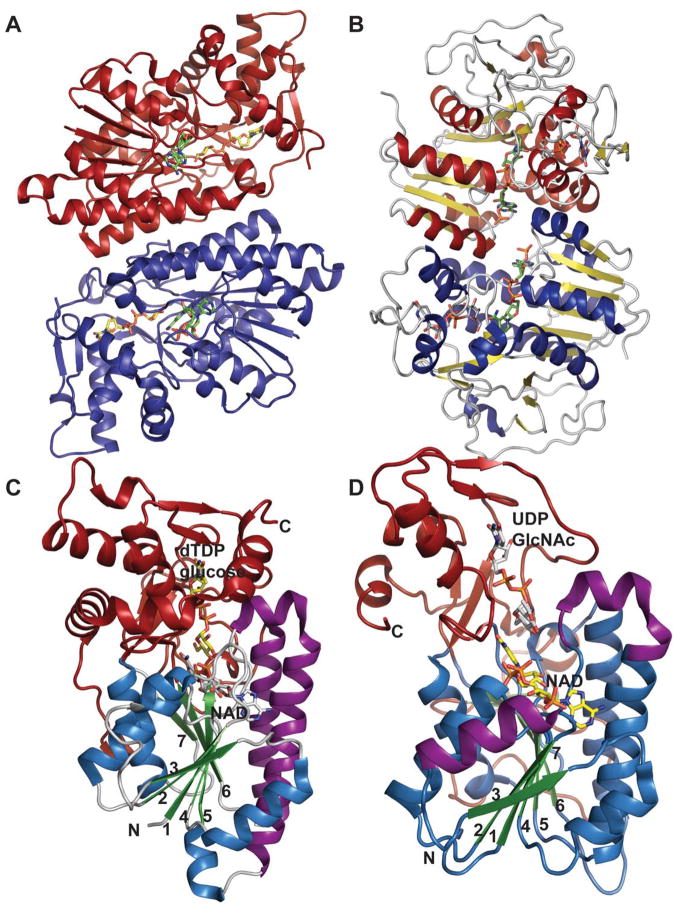
Representative global folds and monomeric structures of retaining and inverting dehydratases in the SDR structural family. Dimer (A) and monomer (C) of the retaining dehydratase RmlB bound to dTDP-Glucose and NAD from Salmonella enterica (PDB 1KEU). Dimer (B) and monomer (D) of the inverting dehydratase FlaA1 from Helicobacter pylori bound to UDP-GlcNAc and NADPH (PDB 2GN6). The core Rossmann fold is colored in blue/green, C-terminal domain highlighted in red, and purple signifies the helices involved in dimerization.
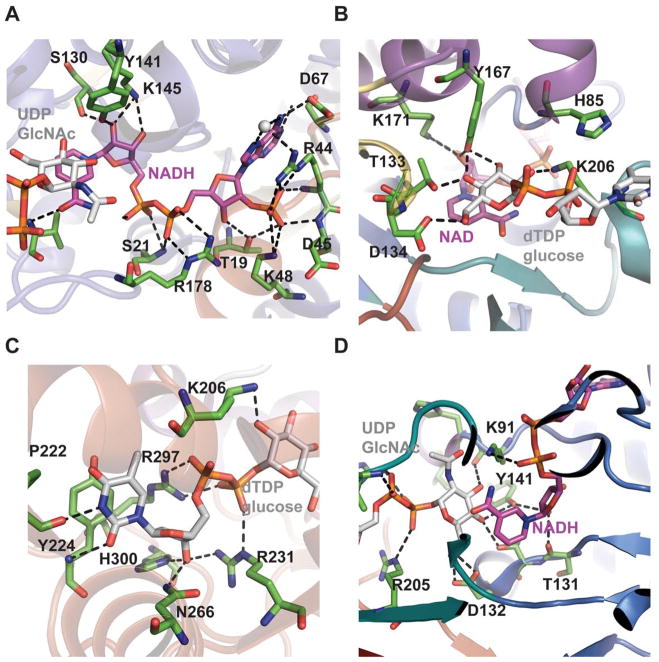
SDR structural family dehydratase active-site interactions. The cofactor binding sites of Helicobacter pylori FlaA1 bound to UDP-GlcNAc and NADP (PDB 2GN6) and Salmonella enterica RmlB bound to NAD+ and dTDP-glucose (PDB 1KEU) are highlighted in panels (A) and (B), respectively. The corresponding sugar binding sites of FlaA1 and RmlB are highlighted in panels (C) and (D), respectively.
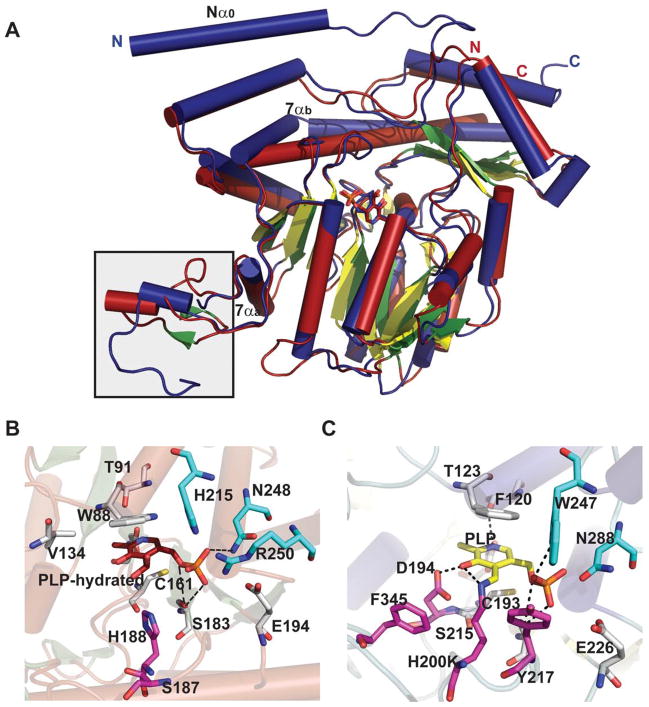
AATstructural family dehydrase global fold and active-site interactions. A. Overlay of ColD from Escherichia coli bound to hydrated PLP (red, PDB 2GMS) and E1 (H220K) from Yersinia pseudotuberculosis bound to PLP (blue, PDB 3BB8). The boxed flexible loop indicates the region that binds to the E1 [2Fe-2S] cluster. B. The active sites of ColD and E1 are represented in panels B and C, respectively. Letters ‘C’ and ‘N’ represent C and N-terminus, respectively.
Global fold of epimerases belonging to cupin structural family. A. dTDP-4-dehydrorhamnose 3,5-epimerase RmlC from Pseudomonas aeruginosa bound to dTDP-4-keto-rhamnose (PDB 2IXK ). N-terminal domain colored purple, C-terminal domain colored blue, core-domain in yellow and green B.
Amycolatopsis orientalis dTDP-3-amino-4-keto-2,3,6-trideoxy-3-C-methyl-glucose-5-epimerase EvaD from (PDB 1OFN). C.
Aneurinibacillus thermoaerophilus dTDP-4-keto-6-deoxy-D-glucose-3,4-ketoisomerase FdtA bound to dTDP (PDB 2PA7). In panels B and C, red/green represents one monomer while blue/yellow represents another.
Global fold of epimerases belonging to SDR structural family. A. Tetramer of CDP-tyvelose-2-epimerase from Salmonella enterica subsp. enterica serovar Typhi bound to CDP and NAD (PDB 1ORR). B. ADP-L-glycero-D-mannoheptose-6-epimerase from Escherichia coli (PDB 1EQ2). C. UDP-galactose epimerase from Escherichia coli bound to UDP-Glc and NAD (PDB 1XEL). For panels A and C, each monomer is represented by a distinct color. In panel B the Rossmann fold is highlighted in blue/green and the C-terminal domain in red/yellow.
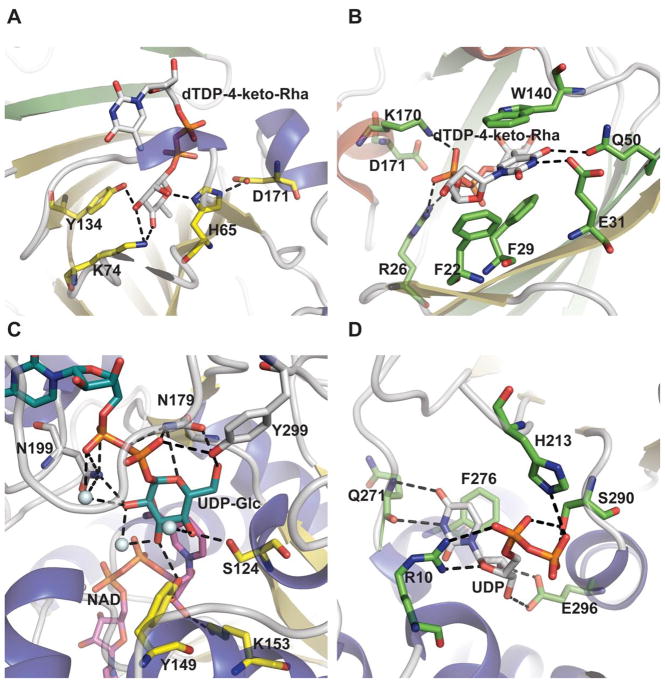
Epimerase active site architecture. A. Interaction of Pseudomonas aeruginosa dTDP-4-dehydrorhamnose 3,5-epimerase RmlC bound to dTDP-4-keto-rhamnose (PDB 2IXK ). B. Nucleotide base interactions of the RmlC ligand-bound complex. C. Interaction of sugar within Escherichia coli UDP-galactose-4-epimerase bound to UDP-Glc and NAD (PDB 1XEL). D.
Escherichia coli UDP-N-acetylglucosamine 2-epimerase bound to UDP (PDB 1F6D).
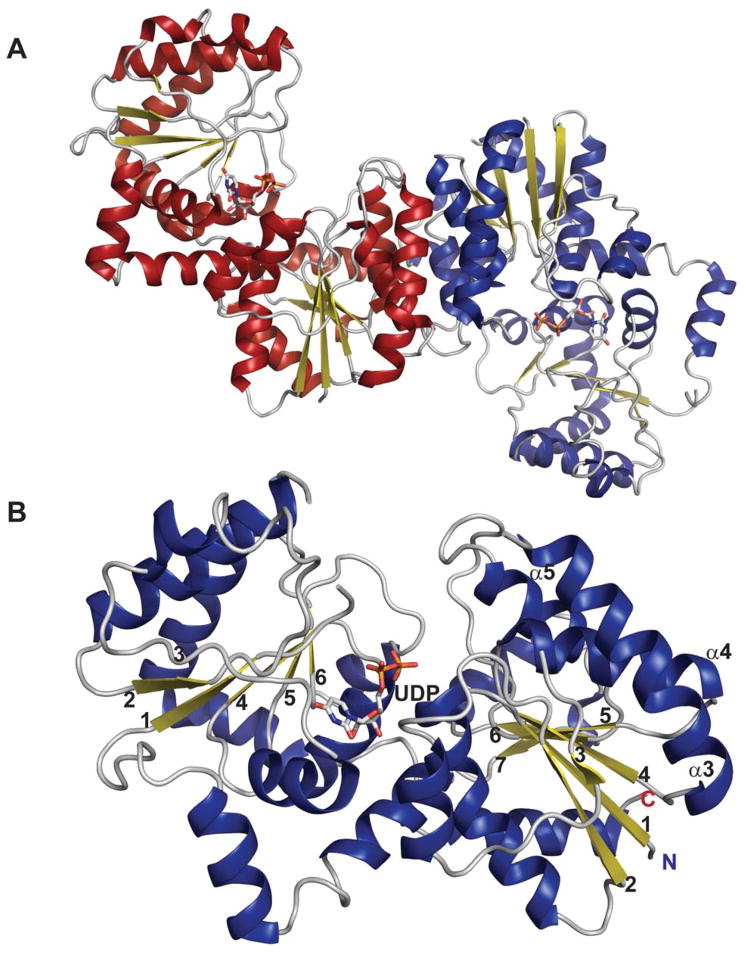
Global fold and monomeric structure of epimerases belonging to the GT-B structural family. Dimeric (A) and monomeric (B) structure of Escherichia coli UDP-N-acetylglucosamine 2-epimerase bound to UDP (PDB 1F6D). The labeled secondary structure elements (α3, α4, α5) signify the helices involved in dimerization.
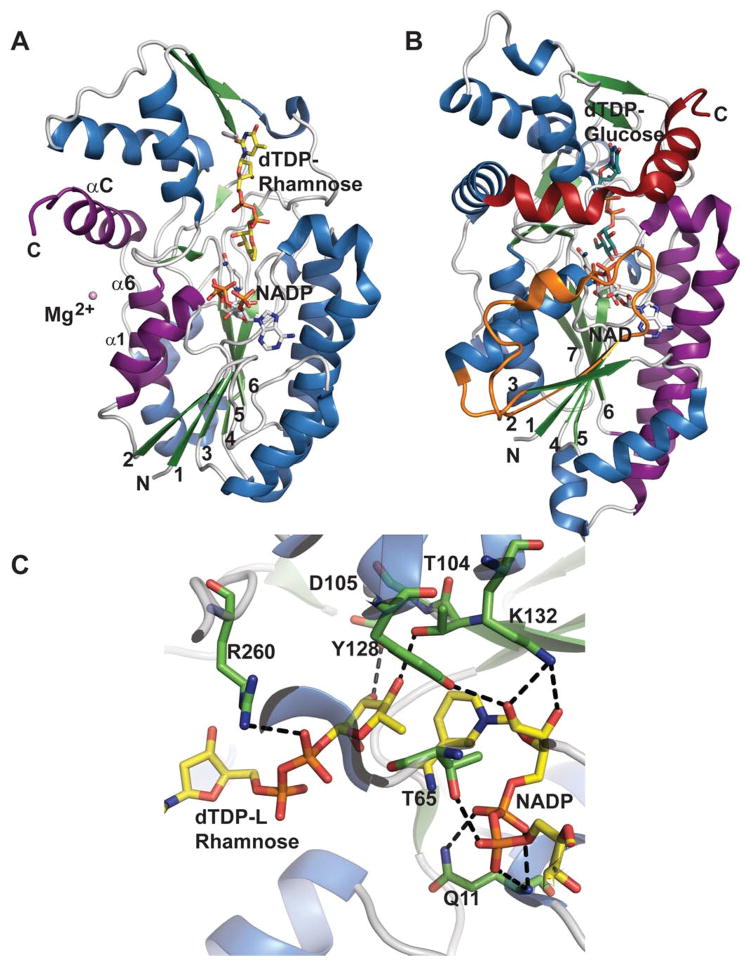
Comparison of sugar ketoreductase and sugar 4,6-dehydratase SDR folds. A.
Salmonella enterica RmlD bound to dTDP-Rhamnose and NADPH (PDB 1KC3). B.
Salmonella enterica RmlB bound to dTDP-Glucose and NAD+ (PDB 1KEU). Orange represents additional RmlB secondary structure (β2-L2-α2), red distinguishes an extra RmlB C-terminus helix and purple signifies secondary structures involved in dimerization C. The active site of RmlD.
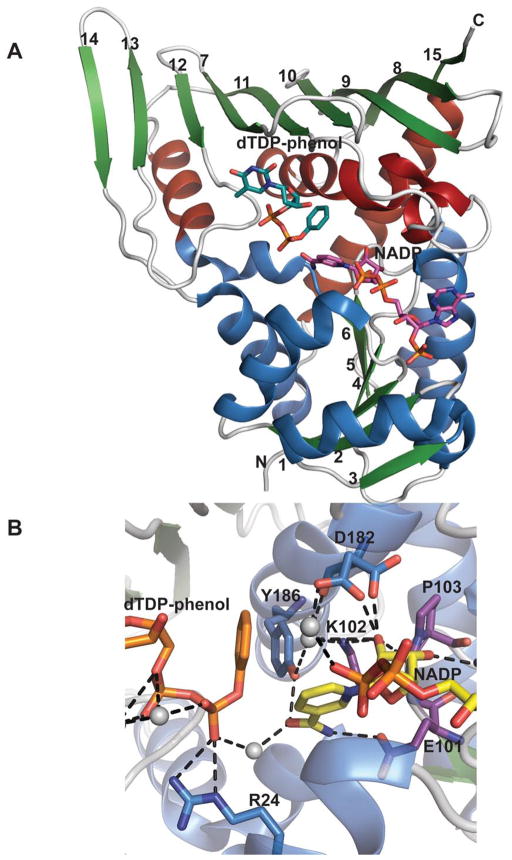
Monomeric structure of an epimerase belonging to the GFOR structural family. A. Monomeric structure of Actinomadura kijaniata KijD10 from (PDB 3RC1) bound to NADP and dTDP-phenol. Helices within the N- and C- terminal domains are represented by distinct colors. B. KijD10 active site. Purple signifies residues within the conserved EKP motif, grey spheres highlight water molecules, and the side chain of Asp182 adopts two different configurations.
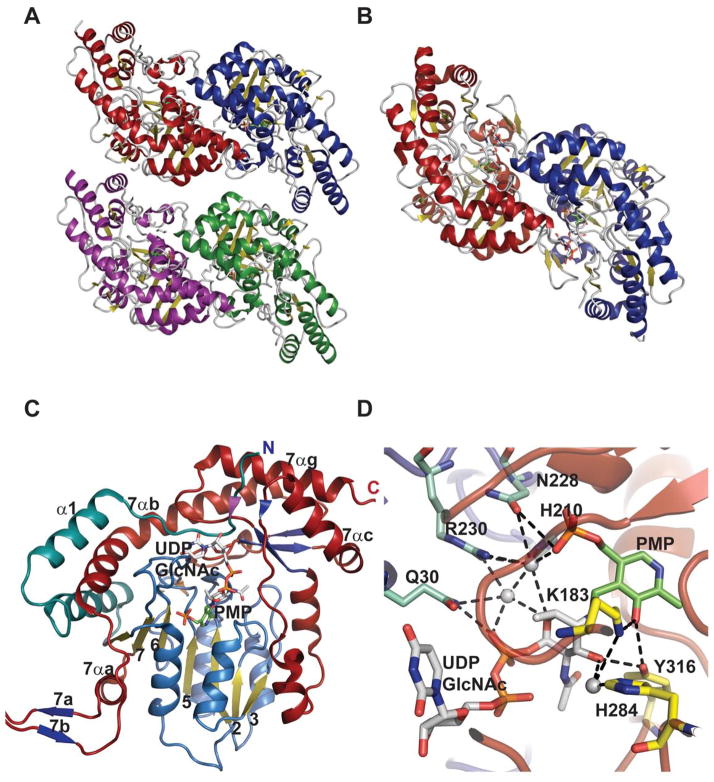
Representative global folds and monomeric structures for sugar aminotransferases belonging to the AAT family. A. Tetramer of Streptomyces venezuelae DesV, a 3-aminotransferase (PDB 2OGA). B. Dimer of Streptomyces venezuelae DesI, a 4-aminotransferase (PDB 2PO3). In panels A and B, each monomer is represented by a distinct color. C. Monomer of Helicobacter pylori PseC bound to UDP-GlcNAc and PMP (PDB 2FNU). D. Active site of PseC bound to UDP-GlcNAc and PMP.
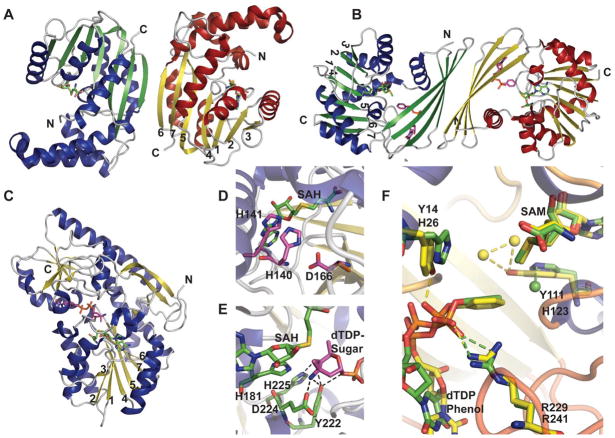
Structure and active site architecture of sugar methyltransferases. A. Dimer of Lechevalieria aerocolonigenes RebM, a rebeccamycin sugar O-methyltransferase (PDB 3BUS). B. Dimer of Streptomyces venezuelae DesVI a N,N,-dimethyltransferase (PDB 3BXO). C.
Micromonospora chalcea TcaB9, a C-methyltransferase (PDB 3NDJ) involved in the biosynthesis of D-tetronitrose. D. Active site of RebM. E. Active site of TcaB9 bound to dTDP-3-amino-2,3,6-trideoxy-4-keto-3-methyl-D-glucose. F. Overlay of the active sites of N-methyltransferases TylM1 (green, PDB 3PFG) and DesVI (yellow). Spheres represent water molecules.
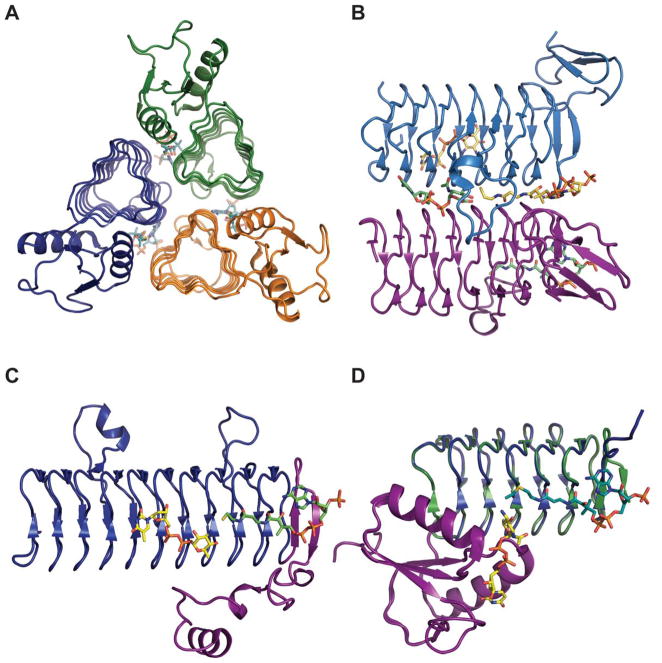
Representative global folds and monomeric structures for sugar N-acyltransferases belonging to the LβH family. A. Trimer of Campylobacter jejuni PglD (PDB 3BSW). B. Dimer interface of Bordetella petrii WlbB with subunit A and B substrates colored yellow and green, respectively (PDB 3MQH). C. Monomer of Thermoanaerobacterium thermosaccharolyticum QdtC bound to CoA and dTDP-3,6-dideoxy-3-amino galactose (PDB 3FSC). D. Overlay of monomer of PglD ligand-bound complexes - UDP-2-acetamido-4-amino-6-deoxy-glucose-bound (PDB 3BSS) and AcCoA-bound (PDB 3BSY). In panels C and D, the N-terminal domains are colored purple.
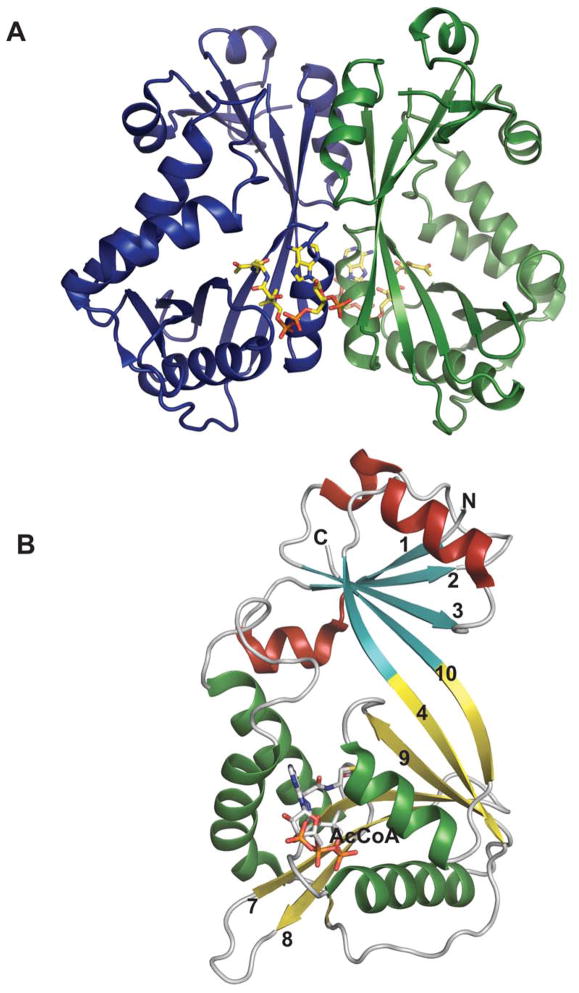
Representative global fold and monomeric structure for a sugar N-acytransferase belonging to the GNAT family. Dimeric (A) and monomeric (B) structure of Escherichia coli WecD from bound to acetyl-CoA (PDB 2FT0).
Structures of N-oxidases. A. Tetramer of Micromonospora carbonacea var. africana nitrososynthase EvdC (PDB 3MXL). B. dTDP binding site of Actinomadura kijaniata nitrosynthase KijD3 (PDB 3M9V). C. Monomer of KijD3. Green color distinguishes N-terminal α-helical domain, β-sheet domain is colored yellow, and purple highlights the C-terminal α-helical domain which is also the tetramerization interface.
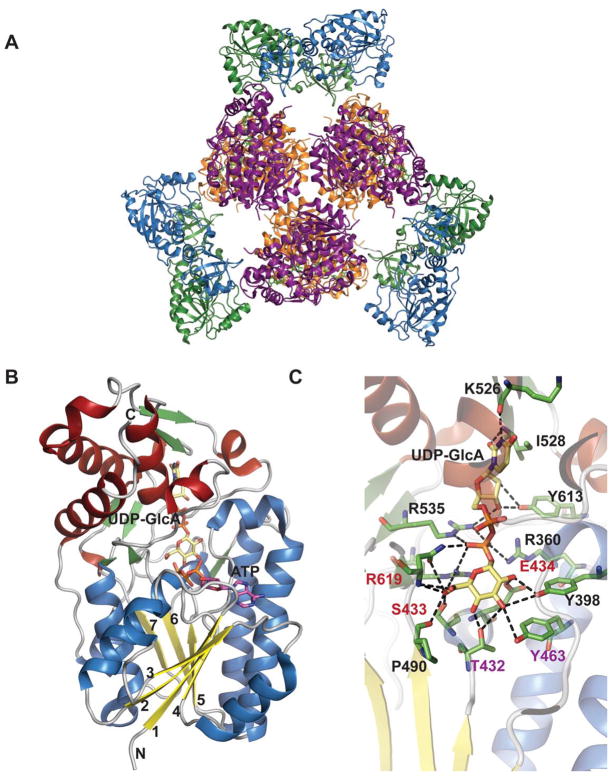
Global fold and dehydrogenase domain of E. coli ArnA. A. Hexameric structure of full-length ArnA bound to ATP and UDP-GlcA (PDB 1Z7E). B. Dehydrogenase (DH) domain of ArnA. C. Active site of DH domain of ArnA. The residues colored red and purple play a role in catalysis.
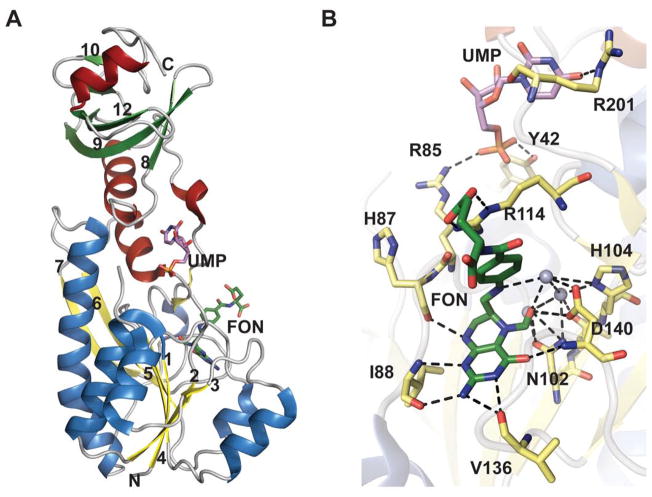
The formyltransferase domain of E. coli ArnA. A. ArnA formyltransferase domain bound to UMP and N-5-formyltetrahydrofolate (FON) (PDB 2BLN). Helices in N- and C-terminal domains are colored differently. B. Active site of formyltransferase domain bound to UMP and N-5-Formyltetrahydrofolate. Grey spheres are water molecules.
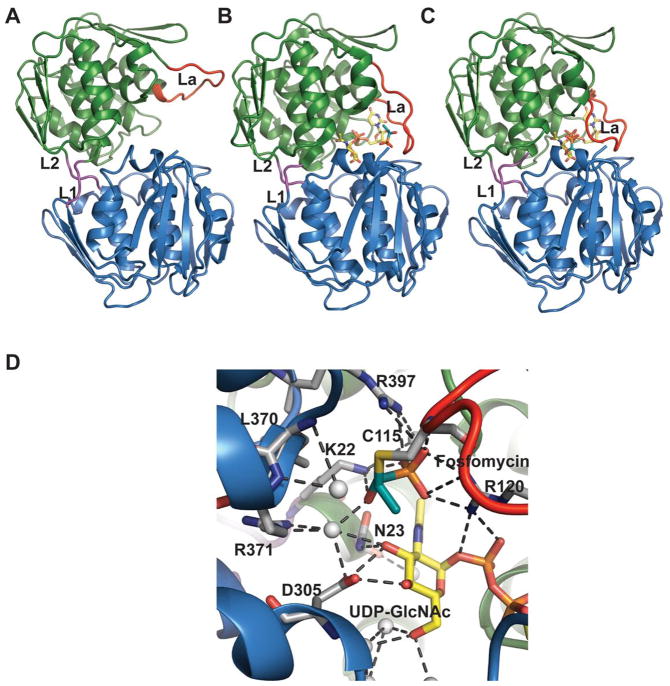
Structures of MurA where the active site flexible loop is colored red and domain linkers are colored magenta. A. Open, ligand-free form of E. cloacae MurA (PDB 1NAW). B. Half open conformation of H. influenzae MurA complexed with UDP-GlcNAc and fosfomycin (PDB 2RL2). C. Closed conformation of E. coli MurA complexed with UDP-GlcNAc and fosfomycin (PDB 1UAE). L1 and L2 are the two linker regions, loop La is an active site loop containing residues P111-P121. D. Active-site interactions within E. coli MurA complexed with UDP-GlcNAc and fosfomycin.
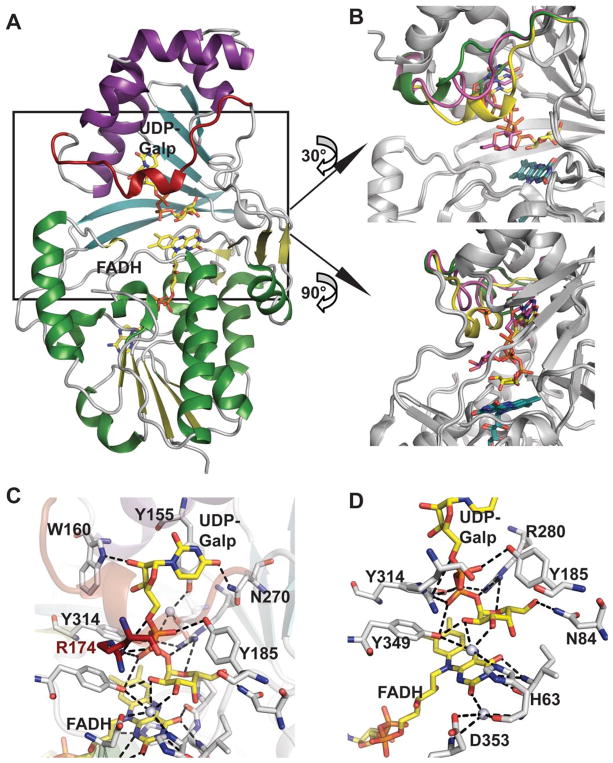
A. Structure of UDP-galactopyranose mutase (UGM)-substrate complex from Klebsiella pneumonia (PDB 3INT). Domains 1 (FAD binding domain), 2 (substrate binding domain) and 3 (dimerization domain) are colored green/yellow, cyan and purple, respectively, the mobile loop is colored red, and substrate (UDP-Galp) and cofactor (FAD) are rendered as sticks and colored yellow. B. Two different views of the conformation of the mobile loop in the superposed structures of UGMs bound to UDP-Glc (green, 3GF4), UDP-Galp in oxidized state (magenta, PDB 3INR) and UDP-Galp in reduced state (yellow, PDB 3INT). C. The substrate binding site highlighting uridine and diphosphate interactions. D. The sugar and flavin binding region. Water molecules are illustrated as spheres.
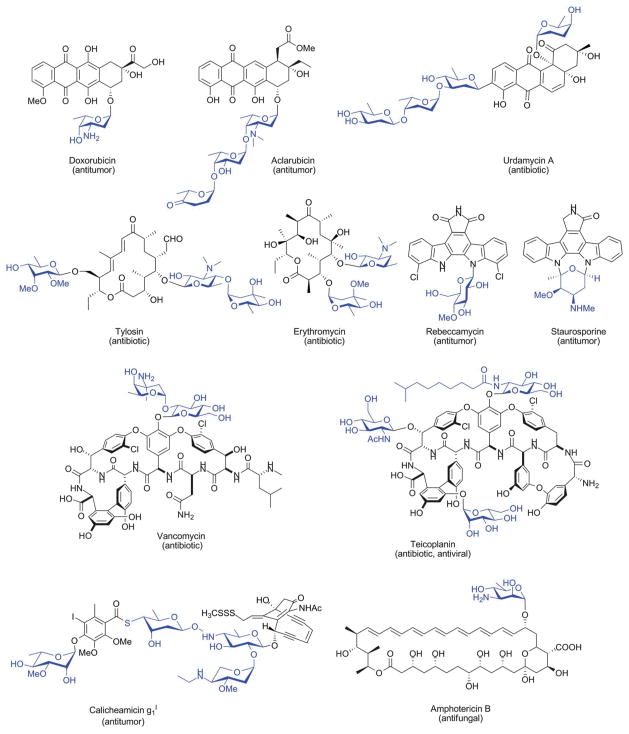
Representative glycosylated natural products of microbial origin. Appended sugars are highlighted in blue.
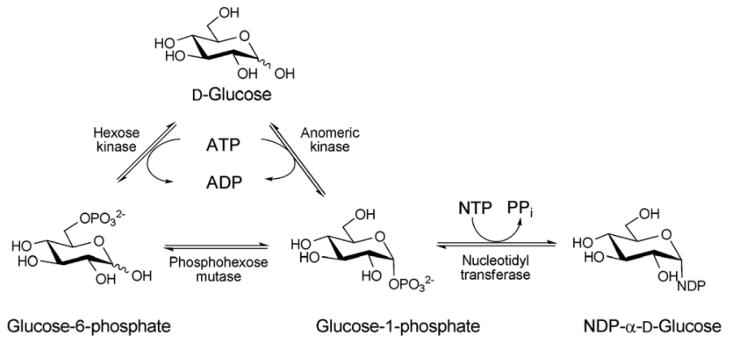
The biosynthesis of NDP-sugars from sugar-1-phosphate. The anomeric kinase reaction presented reflects reactions catalyzed by non-native engineered catalysts.
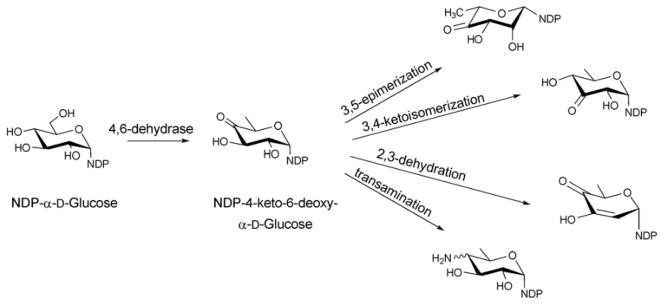
Reactions originating from the common intermediate, NDP-4-keto-6-deoxy-α-D-glucose, which is the product of the 4,6-dehydratase reaction of NDP-D-glucose.
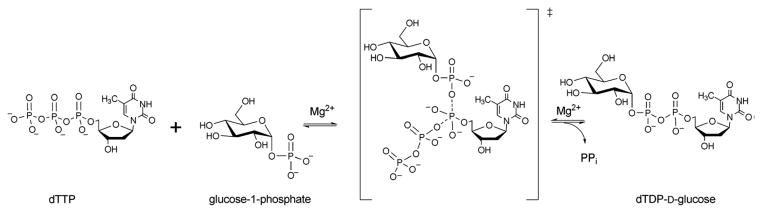
Reaction mechanism of glucose-1-phosphate thymidylytransferase.
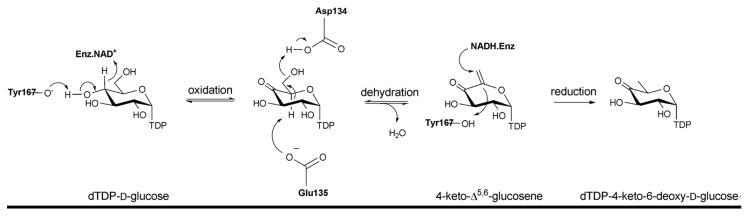
Reaction mechanism of dTDP-glucose-4,6-dehydratase (RmlB).
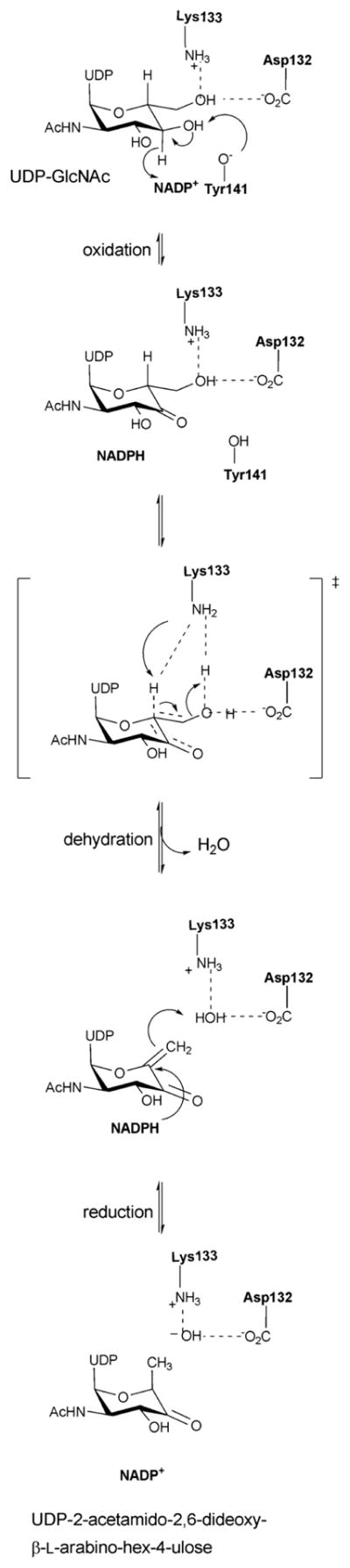
Reaction mechanism of an inverting 4,6-dehydratase (FlaA1).
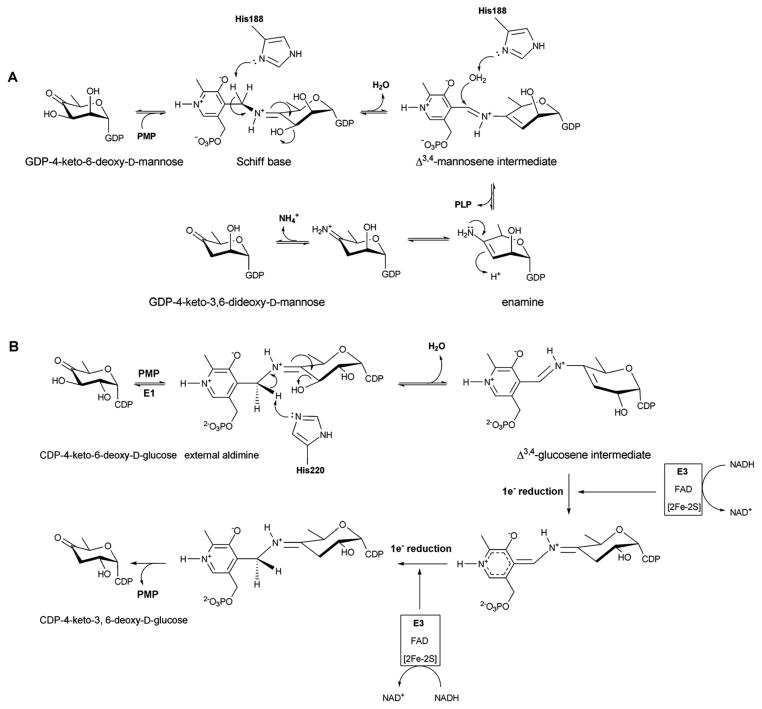
A. Reaction mechanism of GDP-4,6-mannose-dehydratase ColD. B. Reaction mechanism of CDP-6-deoxy-L-threo-D-glycero-4-hexulose 3-dehydrase E1.
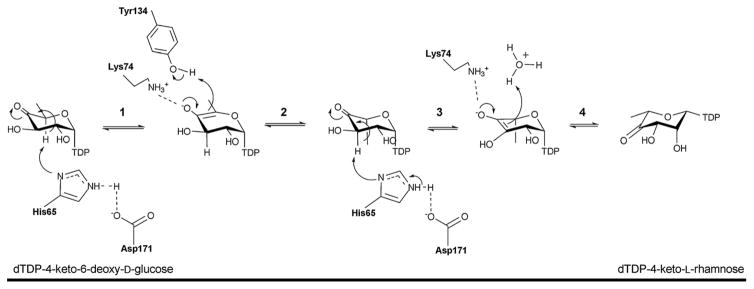
Reaction mechanism of the 3,5-epimerase RmlC. Step 1: C5 proton abstraction by the active site His-Asp diad; step 2: proton addition assisted by active site tyrosine and subsequent inversion; step 3: C3 proton abstraction by the active site His-Asp diad; step 4: proton addition and inversion.
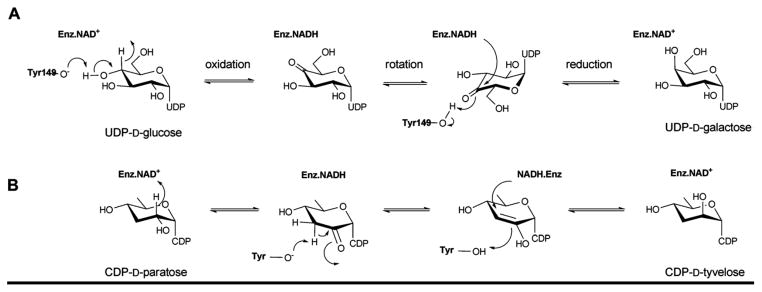
Reaction mechanism employed by UDP-galactose-4-epimerase, which catalyzes the interconversion of UDP-glucose into UDP-galactose (A), and CDP-tyvelose-2-epimerase, which interconverts CDP-tyvelose and CDP-paratose (B).
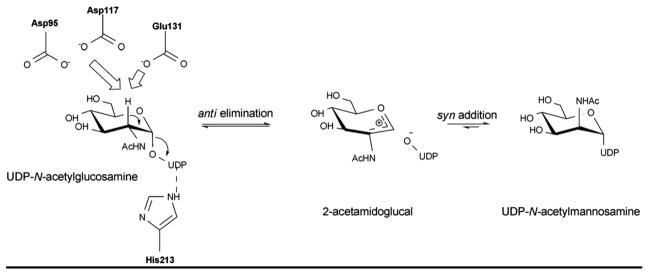
Reaction mechanism of UDP-GlcNAc-2-epimerase, the enzyme that converts UDP-GlcNAc to UDP-ManNAc.
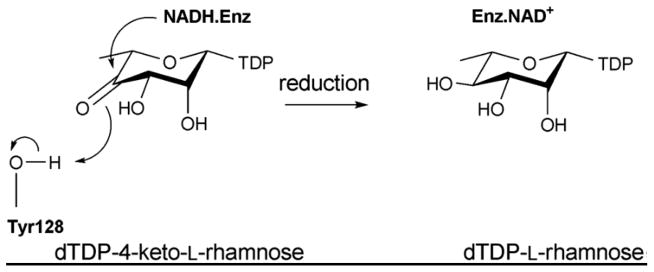
The mechanism of C4 reductase RmlD.
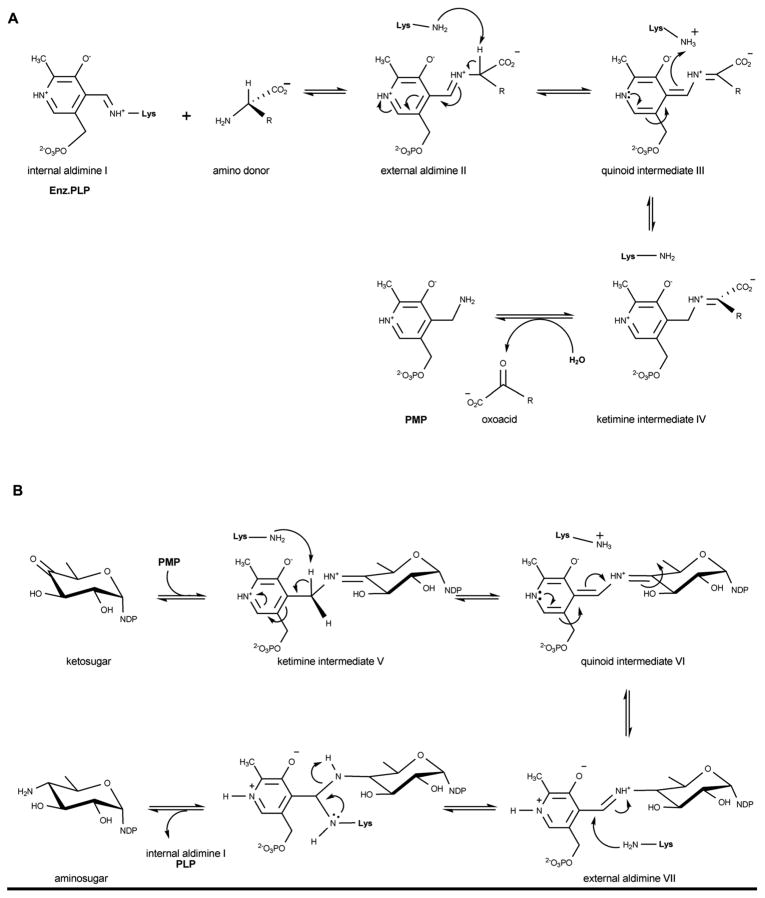
Reaction mechanism employed by aminotransferases. The first half of the reaction enables ‘amine loading’ (namely conversion of PLP to PMP) (A) while the second half of the reaction completes the cycle (B) to ultimately produce an amino sugar and regenerated PLP (bound as internal aldimine).
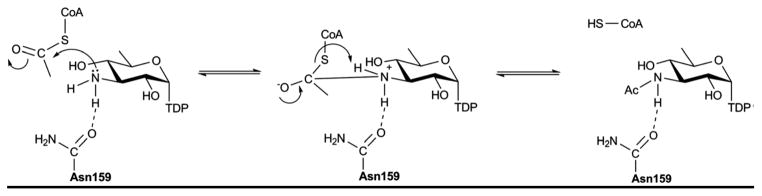
Proposed mechanism for the N-acetyltransferase QdtC.
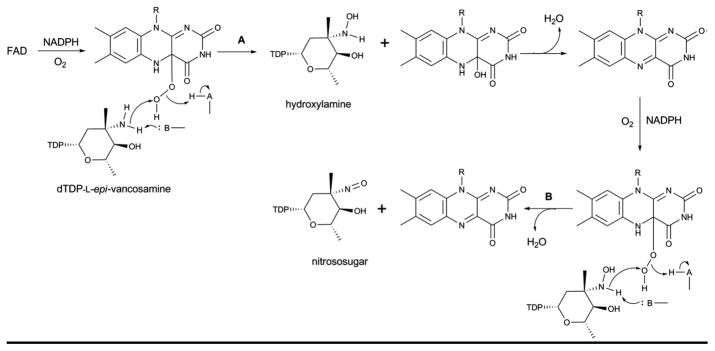
Proposed mechanism for FMO amine oxidase catalysis which provides for sequential oxidation (hydroxylamino – nitroso – nitro) of a target amino sugar.
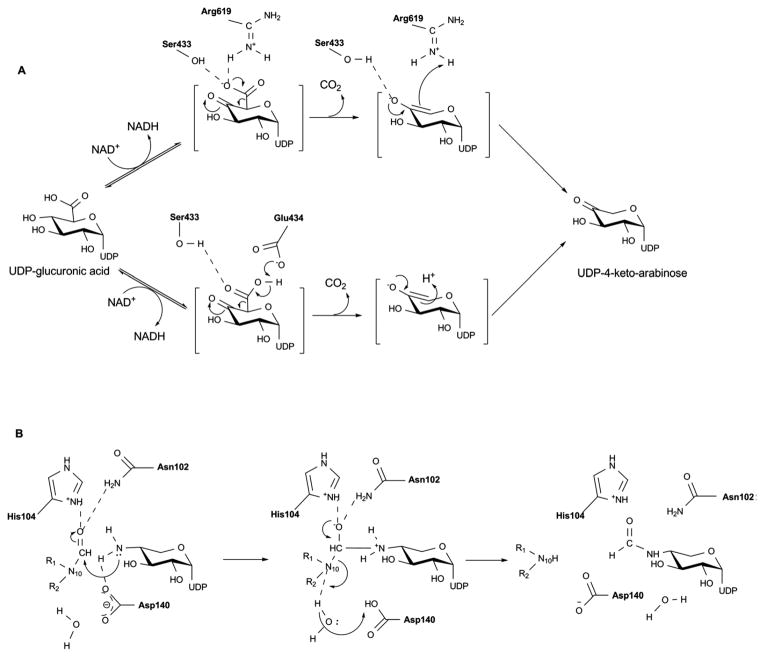
Proposed mechanism for the bifunctional activities of ArnA – dehydrogenase (A) and formyltransferase (B).
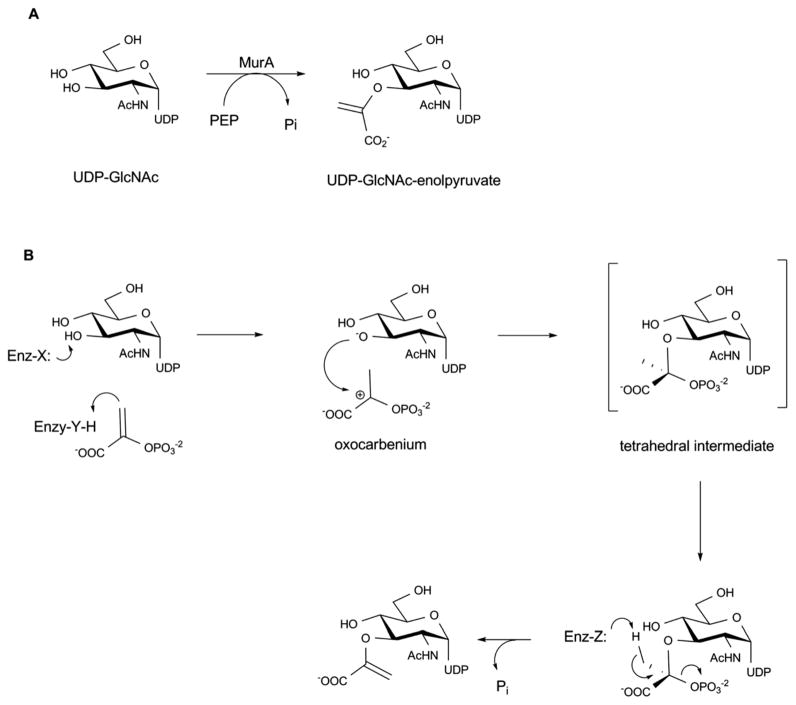
Reaction (A) and proposed mechanism (B) catalyzed by MurA enzyme.
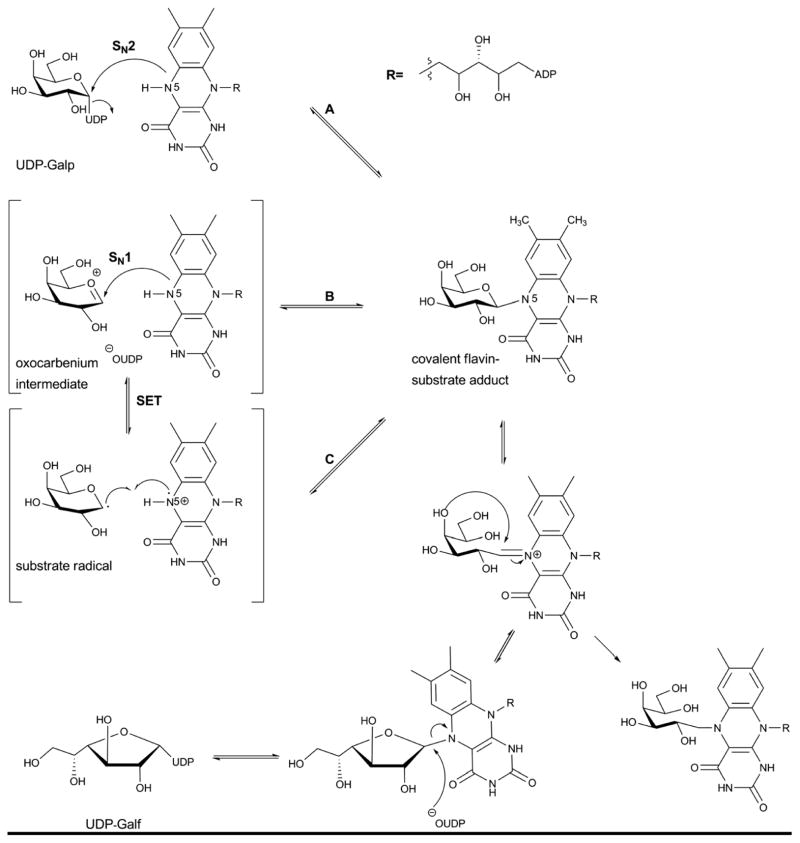
Proposed mechanism of UDP-galactopyranose mutase reaction.
References
-
- Kilcoyne M, Joshi L. Cardiovasc Hematol Agents, Med Chem. 2007;5:186–197. - PubMed
-
- Kren V, Martinkova L. Curr Med Chem. 2001;8:1303–1328. - PubMed
-
- Kren V, Rezanka T. FEMS Microbiol Rev. 2008;32:858–889. - PubMed
-
- Wrodnigg TM, Sprenger FK. Mini Rev Med Chem. 2004;4:437–459. - PubMed
-
- Thorson JS, Hosted TJ, Jiang JQ, Biggins JB, Ahlert J. Curr Org Chem. 2001;5:139–167.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous