

Drosophila lin-52 acts in opposition to repressive components of the Myb-MuvB/dREAM complex
- PMID: 22688510
- PMCID: PMC3434544
- DOI: 10.1128/MCB.00432-12
Drosophila lin-52 acts in opposition to repressive components of the Myb-MuvB/dREAM complex
Abstract
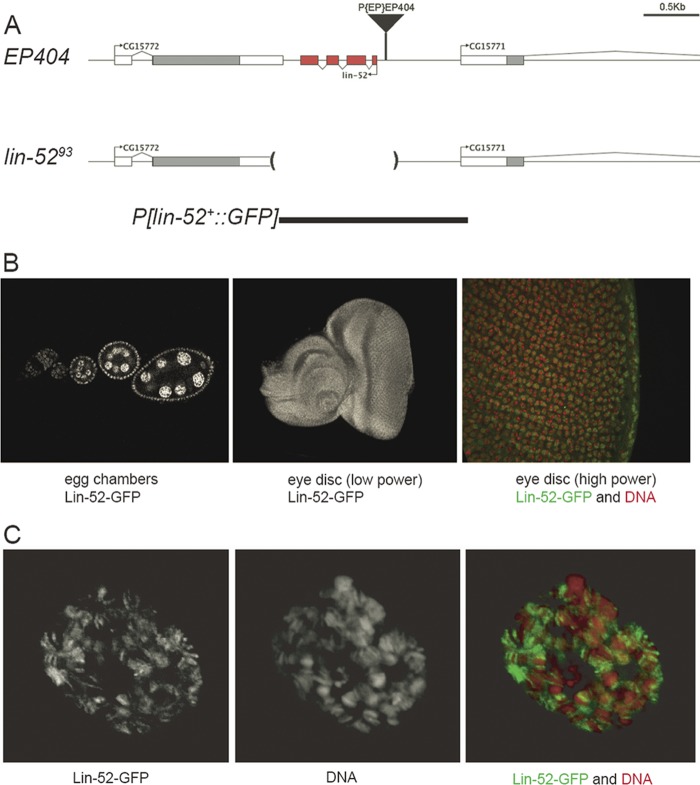
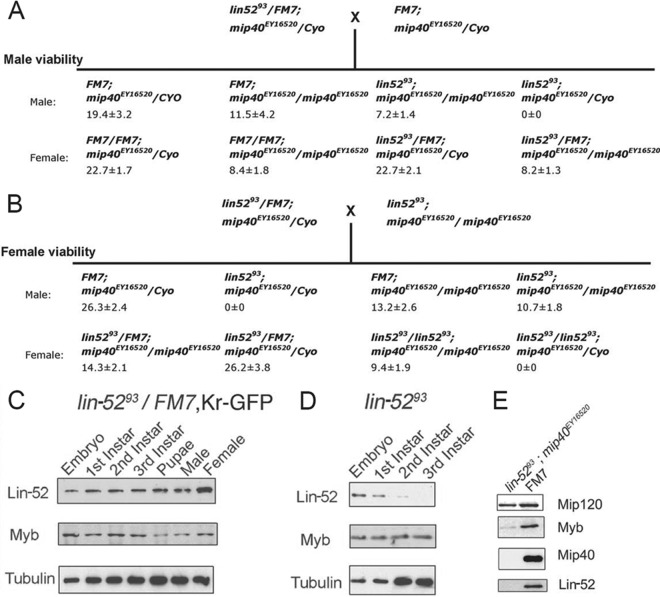
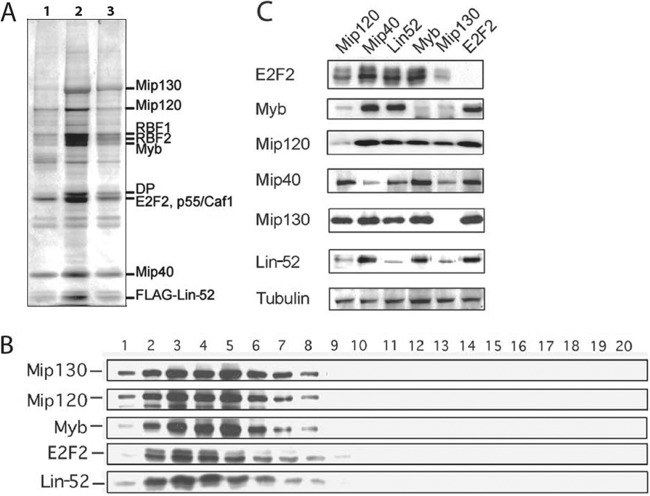
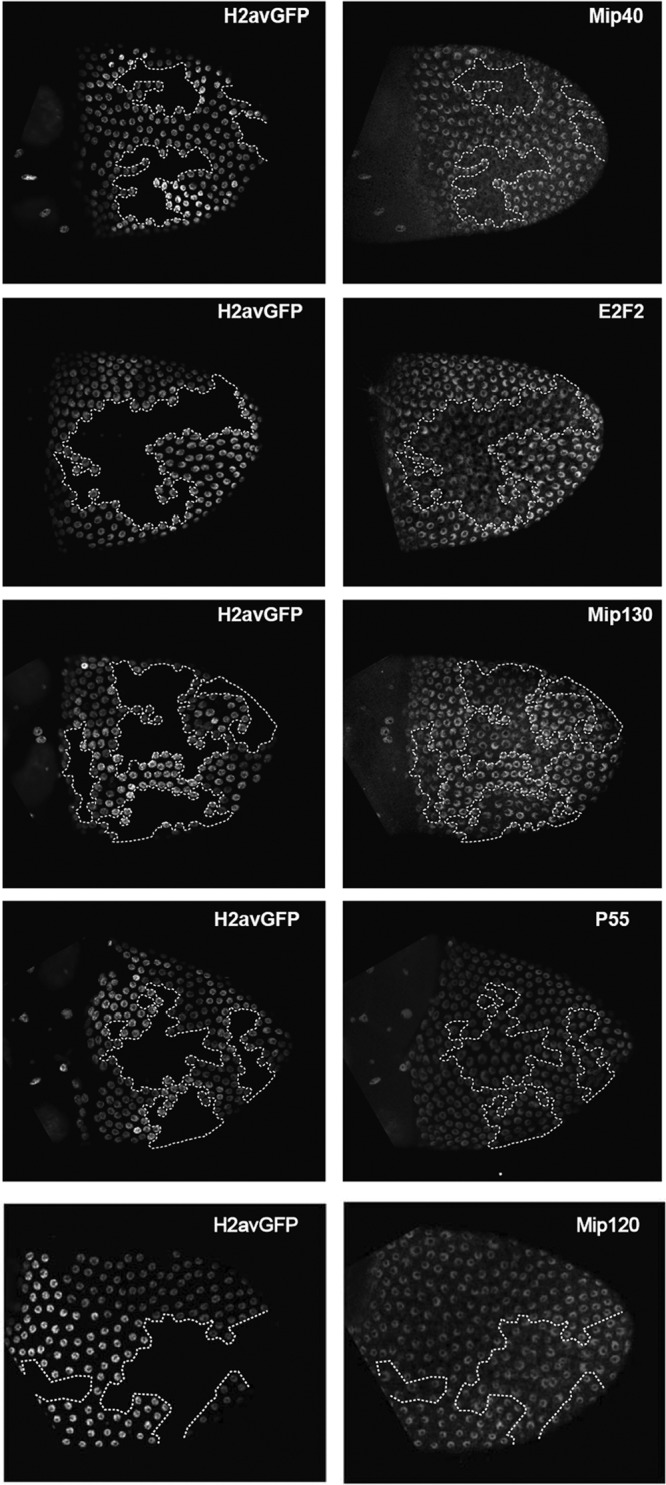
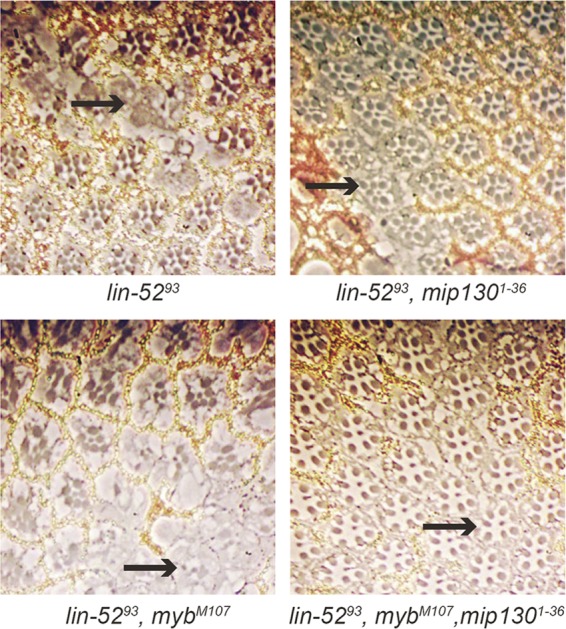
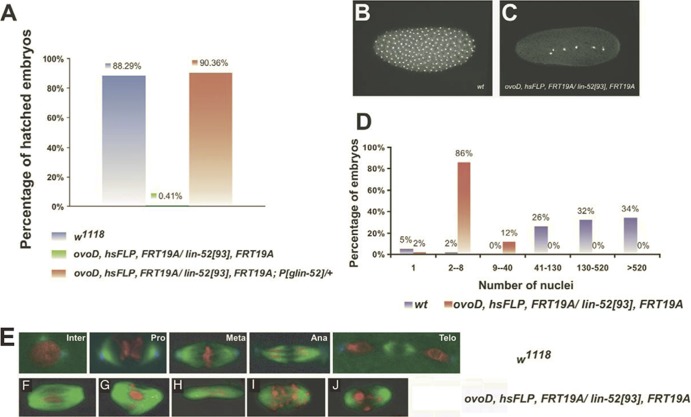
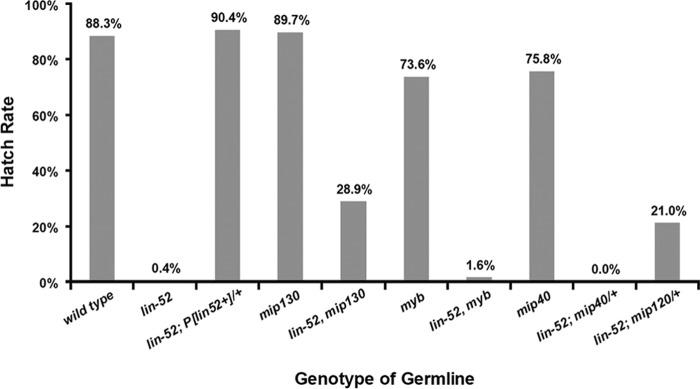
The Drosophila melanogaster Myb-MuvB/dREAM complex (MMB/dREAM) participates in both the activation and repression of developmentally regulated genes and origins of DNA replication. Mutants in MMB subunits exhibit diverse phenotypes, including lethality, eye defects, reduced fecundity, and sterility. Here, we used P-element excision to generate mutations in lin-52, which encodes the smallest subunit of the MMB/dREAM complex. lin-52 is required for viability, as null mutants die prior to pupariation. The generation of somatic and germ line mutant clones indicates that lin-52 is required for adult eye development and for early embryogenesis via maternal effects. Interestingly, the maternal-effect embryonic lethality, larval lethality, and adult eye defects could be suppressed by mutations in other subunits of the MMB/dREAM complex. These results suggest that a partial MMB/dREAM complex is responsible for the lethality and eye defects of lin-52 mutants. Furthermore, these findings support a model in which the Lin-52 and Myb proteins counteract the repressive activities of the other members of the MMB/dREAM complex at specific genomic loci in a developmentally controlled manner.
Figures

References
-
- Beall EL, et al. 2002. Role for a Drosophila Myb-containing protein complex in site-specific DNA replication. Nature 420:833–837 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases