

EphrinA1 is released in three forms from cancer cells by matrix metalloproteases
- PMID: 22688511
- PMCID: PMC3434542
- DOI: 10.1128/MCB.06791-11
EphrinA1 is released in three forms from cancer cells by matrix metalloproteases
Abstract
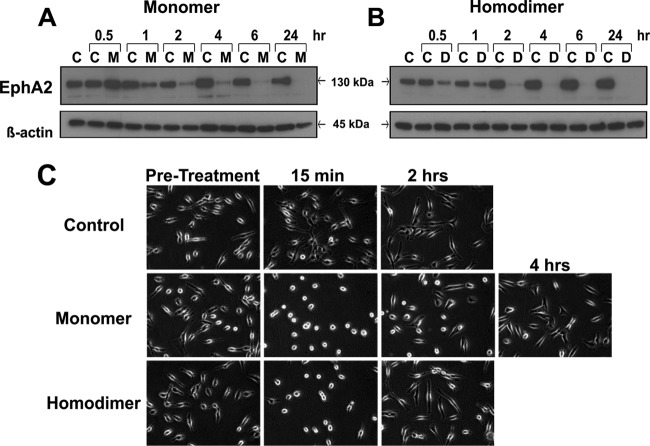
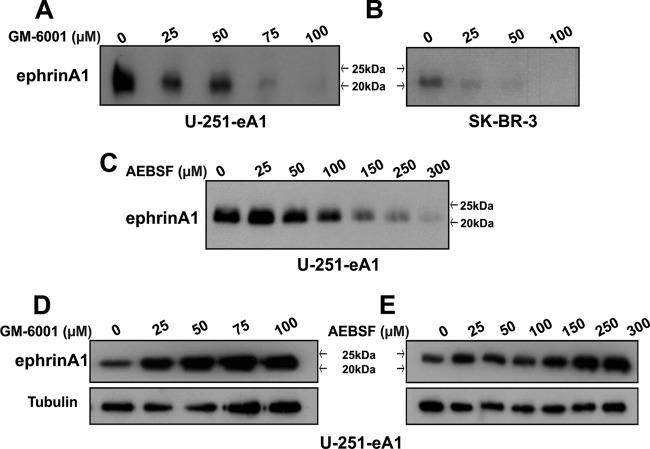
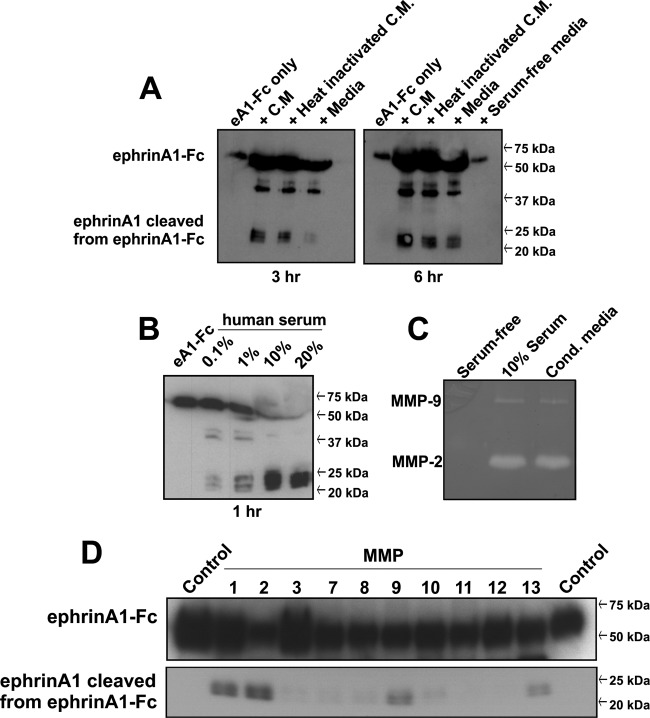
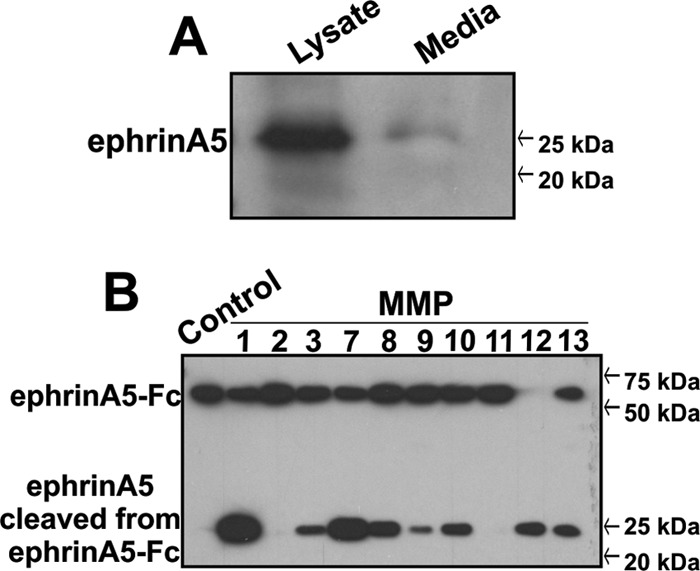
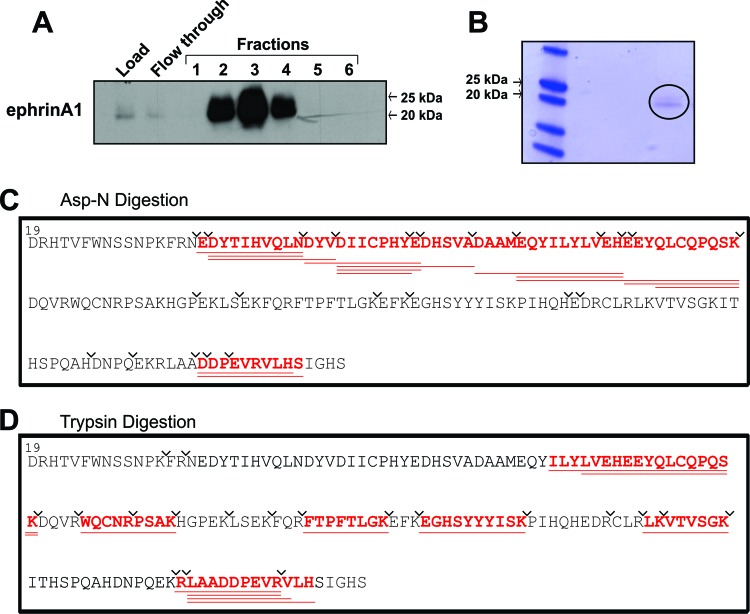
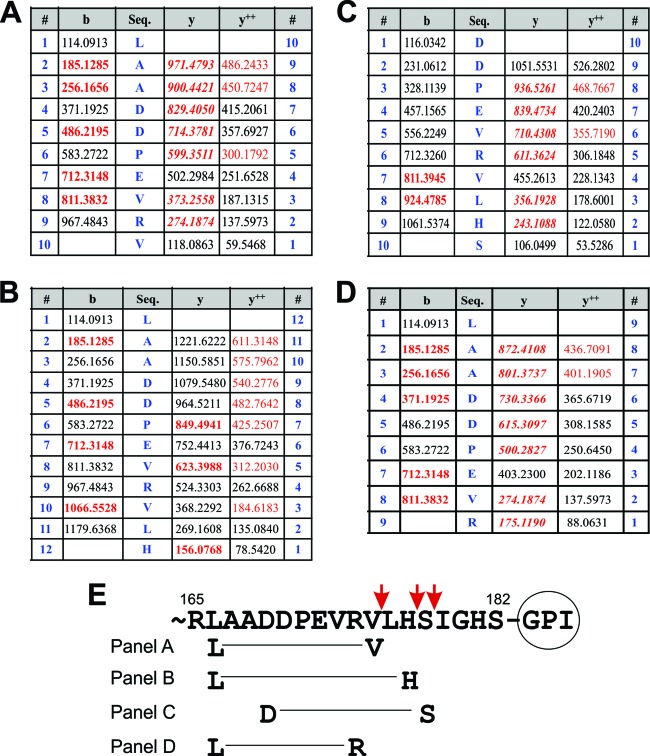
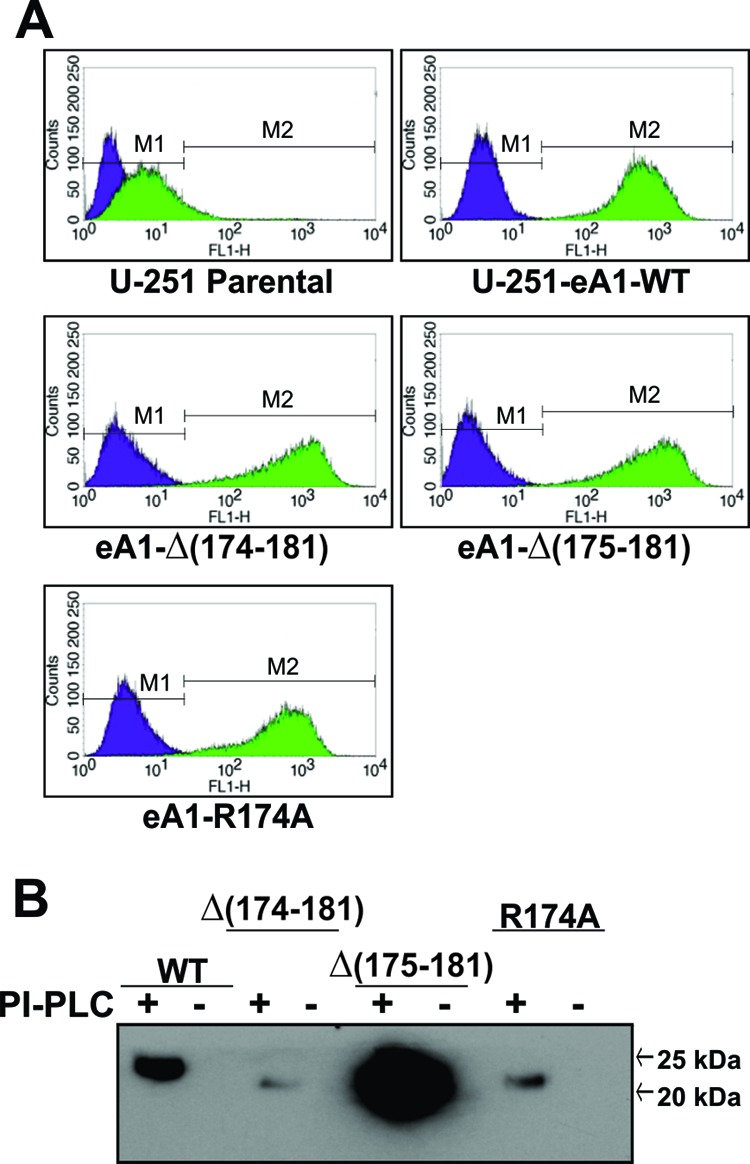
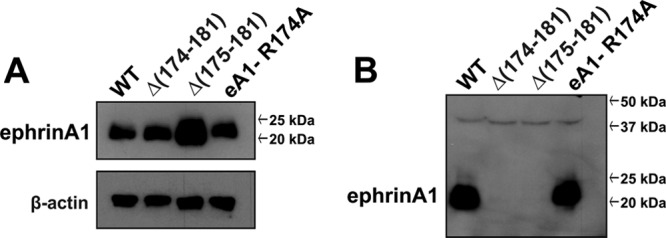
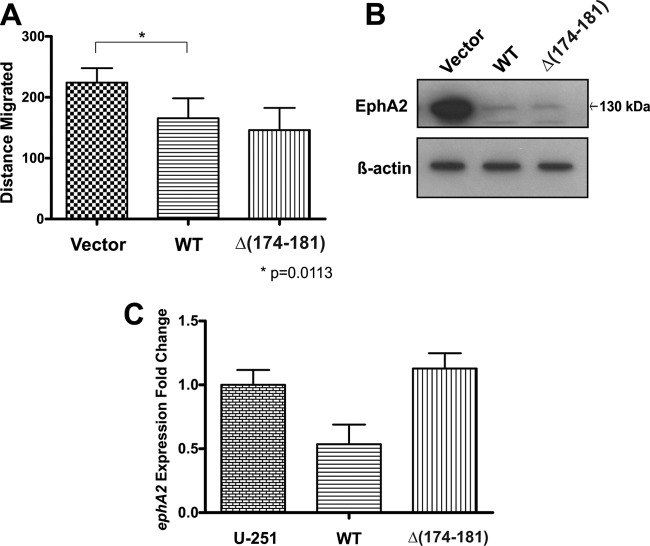
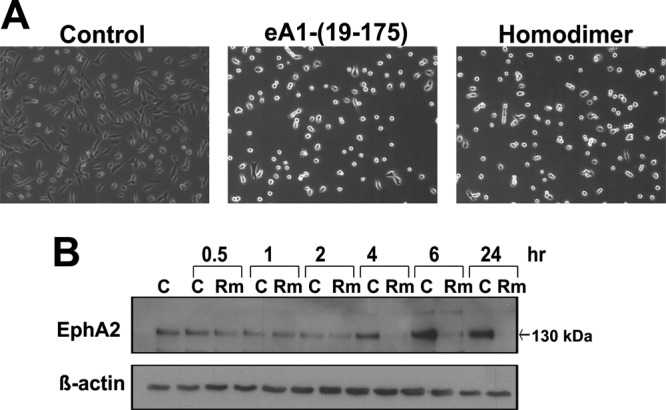
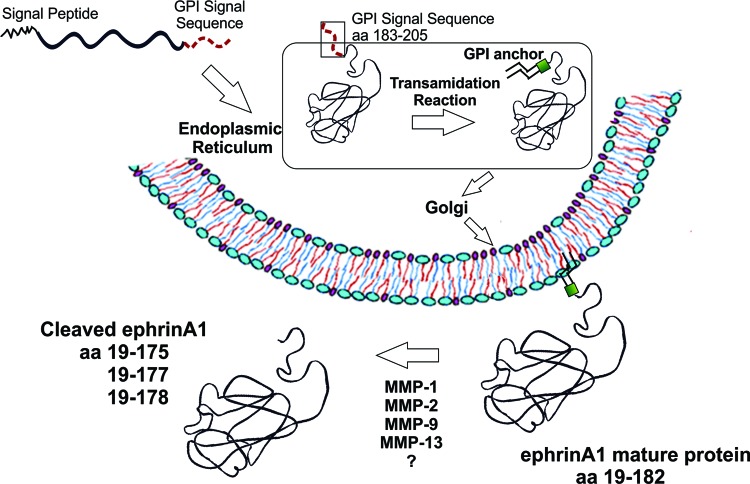
EphrinA1 is a glycosylphosphatidylinositol (GPI)-linked ligand for the EphA2 receptor, which is overexpressed in glioblastoma (GBM), among other cancers. Activation of the receptor by ephrinA1 leads to a suppression of oncogenic properties of GBM cells. We documented that a monomeric functional form of ephrinA1 is released from cancer cells and thus explored the mechanism of ephrinA1 release and the primary protein sequence. We demonstrate here that multiple metalloproteases (MMPs) are able to cleave ephrinA1, most notably MMP-1, -2, -9, and -13. The proteolytic cleavage that releases ephrinA1 occurs at three positions near the C terminus, producing three forms ending in valine-175, histidine-177, or serine-178. Moreover, deletion of amino acids 174 to 181 or 175 to 181 yields ephrinA1 that is still GPI linked but not released by proteolysis, underlining the necessity of amino acids 175 to 181 for release from the membrane. Furthermore, recombinant ephrinA1 ending at residue 175 retains activity toward the EphA2 receptor. These findings suggest a mechanism of release and provide evidence for the existence of several forms of monomeric ephrinA1. Moreover, ephrinA1 should be truncated at a minimum at amino acid 175 in fusions or conjugates with other molecules in order to prevent likely proteolysis within physiological and pathobiological environments.
Figures
Similar articles
-
Soluble monomeric EphrinA1 is released from tumor cells and is a functional ligand for the EphA2 receptor.Oncogene. 2008 Dec 11;27(58):7260-73. doi: 10.1038/onc.2008.328. Epub 2008 Sep 15. Oncogene. 2008. PMID: 18794797 Free PMC article.
-
EphA2 as a novel molecular marker and target in glioblastoma multiforme.Mol Cancer Res. 2005 Oct;3(10):541-51. doi: 10.1158/1541-7786.MCR-05-0056. Mol Cancer Res. 2005. PMID: 16254188
-
EphrinA1-EphA2 interaction-mediated apoptosis and FMS-like tyrosine kinase 3 receptor ligand-induced immunotherapy inhibit tumor growth in a breast cancer mouse model.J Gene Med. 2012 Feb;14(2):77-89. doi: 10.1002/jgm.1649. J Gene Med. 2012. PMID: 22228563 Free PMC article.
-
Up-regulation of EphA2 and down-regulation of EphrinA1 are associated with the aggressive phenotype and poor prognosis of malignant glioma.Tumour Biol. 2010 Oct;31(5):477-88. doi: 10.1007/s13277-010-0060-6. Epub 2010 Jun 23. Tumour Biol. 2010. PMID: 20571968
-
The EphA2 receptor and ephrinA1 ligand in solid tumors: function and therapeutic targeting.Mol Cancer Res. 2008 Dec;6(12):1795-806. doi: 10.1158/1541-7786.MCR-08-0244. Mol Cancer Res. 2008. PMID: 19074825 Free PMC article. Review.
Cited by
-
Simultaneous targeting of Eph receptors in glioblastoma.Oncotarget. 2016 Sep 13;7(37):59860-59876. doi: 10.18632/oncotarget.10978. Oncotarget. 2016. PMID: 27494882 Free PMC article.
-
Eph- and ephrin-dependent mechanisms in tumor and stem cell dynamics.Cell Mol Life Sci. 2014 Oct;71(19):3685-710. doi: 10.1007/s00018-014-1633-0. Epub 2014 May 4. Cell Mol Life Sci. 2014. PMID: 24794629 Free PMC article. Review.
-
Specific detection of soluble EphA2 fragments in blood as a new biomarker for pancreatic cancer.Cell Death Dis. 2017 Oct 26;8(10):e3134. doi: 10.1038/cddis.2017.545. Cell Death Dis. 2017. PMID: 29072678 Free PMC article.
-
Alzheimer's disease-associated P460L variant of EphA1 dysregulates receptor activity and blood-brain barrier function.Alzheimers Dement. 2024 Mar;20(3):2016-2033. doi: 10.1002/alz.13603. Epub 2024 Jan 7. Alzheimers Dement. 2024. PMID: 38184788 Free PMC article.
-
Combined detection of serum EFNA1 and MMP13 as diagnostic biomarker for gastric cancer.Sci Rep. 2024 Jul 10;14(1):15957. doi: 10.1038/s41598-024-65839-y. Sci Rep. 2024. PMID: 38987376 Free PMC article.
References
-
- Alford SC, Bazowski J, Lorimer H, Elowe S, Howard PL. 2007. Tissue transglutaminase clusters soluble A-type ephrins into functionally active high molecular weight oligomers. Exp. Cell Res. 313:4170–4179 - PubMed
-
- Anand M, Van Meter T, Fillmore H. 2011. Epidermal growth factor induces matrix metalloproteinase-1 (MMP-1) expression and invasion in glioma cell lines via the MAPK pathway. J. Neurooncol. 104:679–687 - PubMed
-
- Bartley TD, et al. 1994. B61 is a ligand for the ECK receptor protein-tyrosine kinase. Nature 368:558–560 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous