

How RecBCD enzyme and Chi promote DNA break repair and recombination: a molecular biologist's view
- PMID: 22688812
- PMCID: PMC3372252
- DOI: 10.1128/MMBR.05026-11
How RecBCD enzyme and Chi promote DNA break repair and recombination: a molecular biologist's view
Abstract
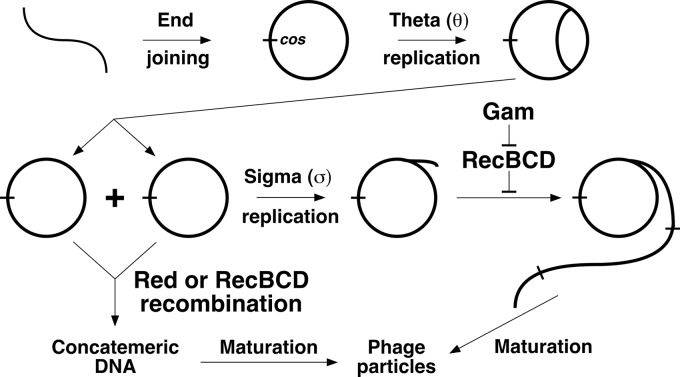
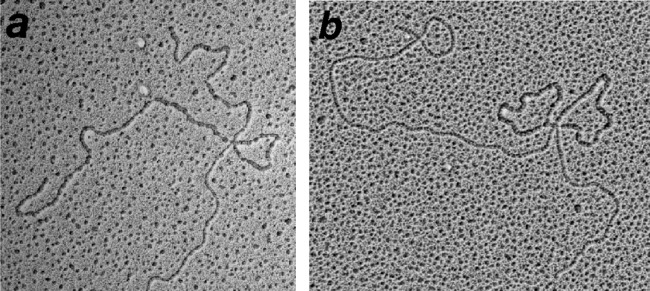
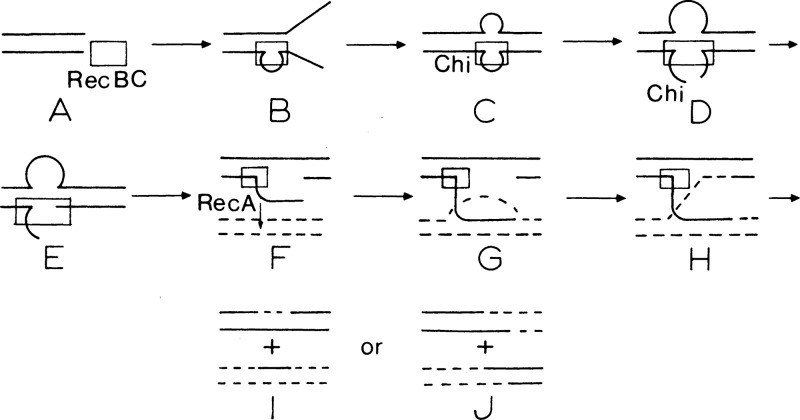
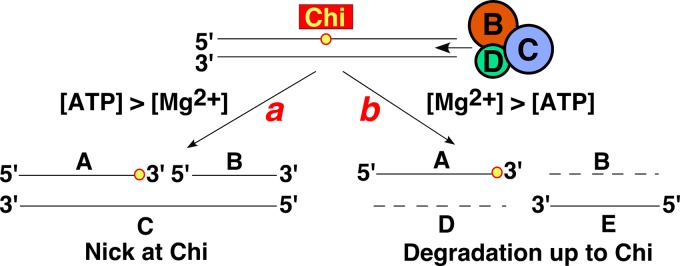
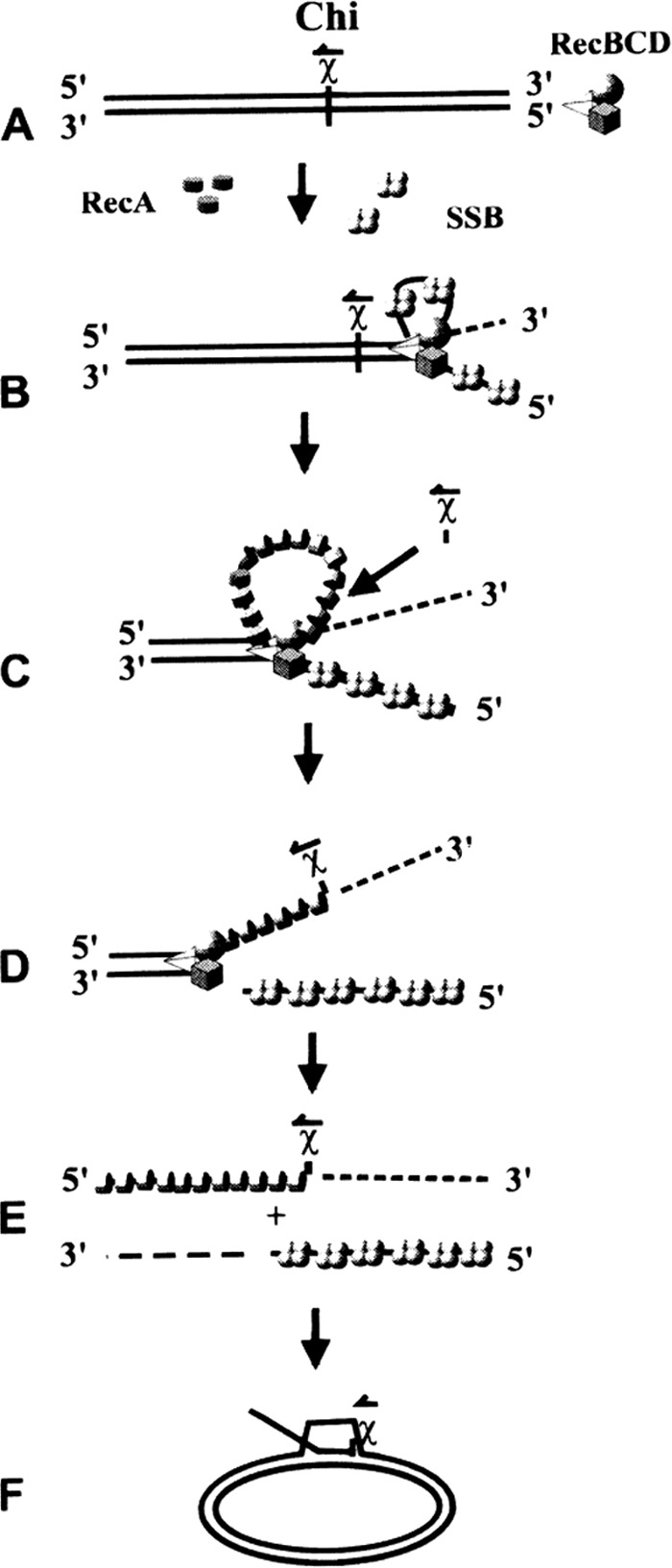
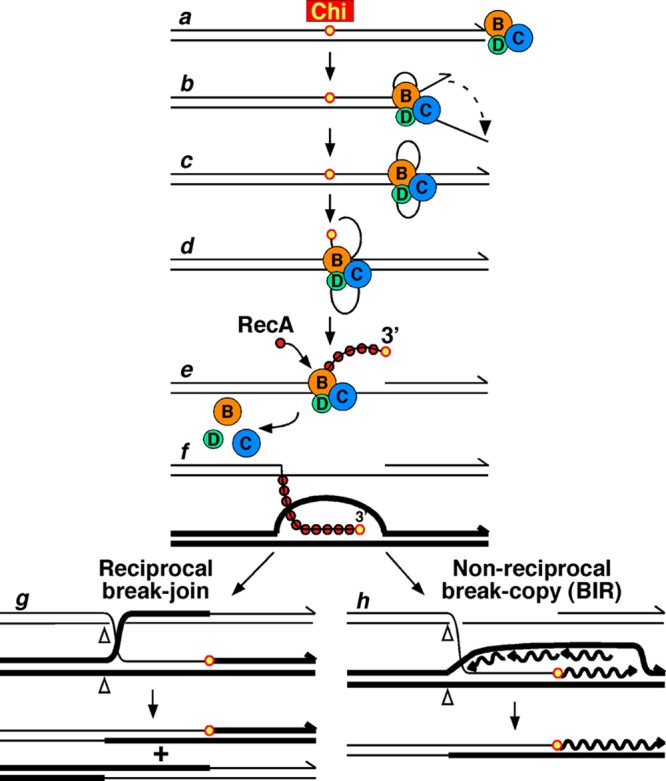
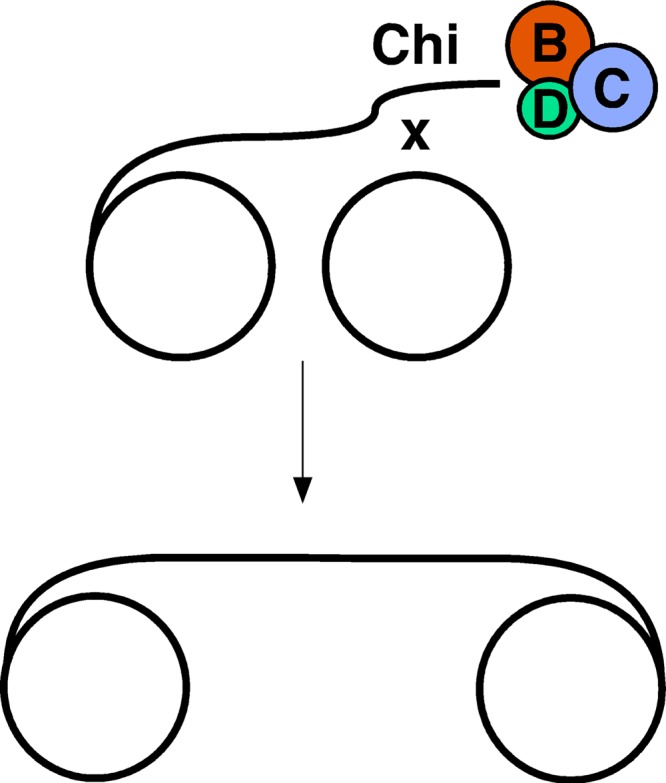
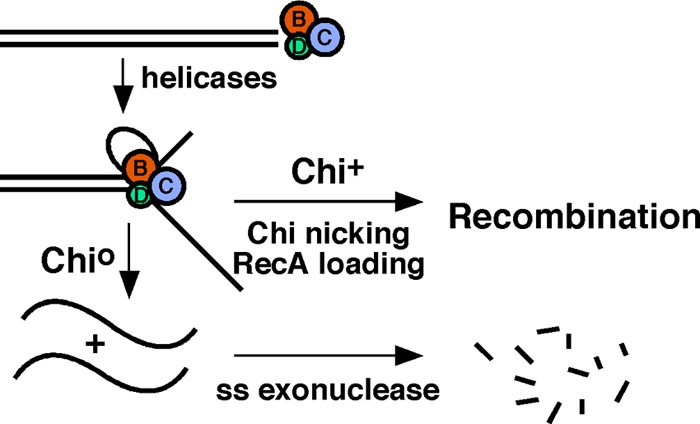
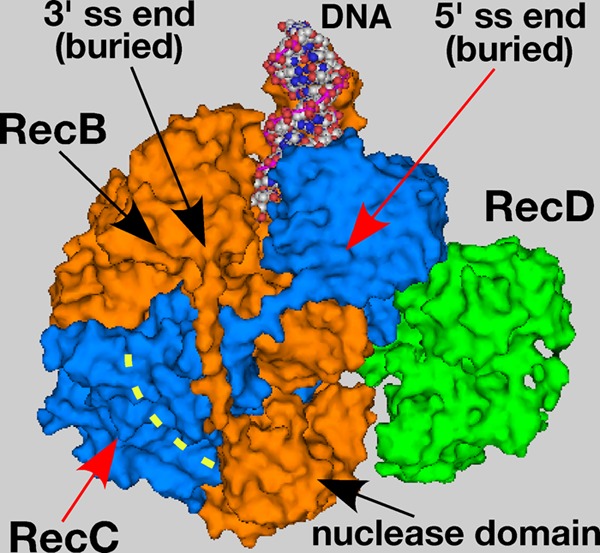
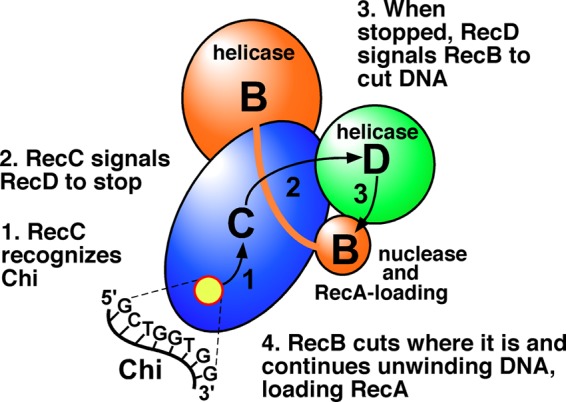
The repair of DNA double-strand breaks (DSBs) is essential for cell viability and important for homologous genetic recombination. In enteric bacteria such as Escherichia coli, the major pathway of DSB repair requires the RecBCD enzyme, a complex helicase-nuclease regulated by a simple unique DNA sequence called Chi. How Chi regulates RecBCD has been extensively studied by both genetics and biochemistry, and two contrasting mechanisms to generate a recombinogenic single-stranded DNA tail have been proposed: the nicking of one DNA strand at Chi versus the switching of degradation from one strand to the other at Chi. Which of these reactions occurs in cells has remained unproven because of the inability to detect intracellular DNA intermediates in bacterial recombination and DNA break repair. Here, I discuss evidence from a combination of genetics and biochemistry indicating that nicking at Chi is the intracellular (in vivo) reaction. This example illustrates the need for both types of analysis (i.e., molecular biology) to uncover the mechanism and control of complex processes in living cells.
Figures


References
-
- Alberts B. 1998. The cell as a collection of protein machines: preparing the next generation of molecular biologists. Cell 92:291–294 - PubMed
-
- Alberts B, et al. 1989. Molecular biology of the cell, 2nd ed Garland Publishing Inc, New York, NY
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
