

Diversity in genetic in vivo methods for protein-protein interaction studies: from the yeast two-hybrid system to the mammalian split-luciferase system
- PMID: 22688816
- PMCID: PMC3372256
- DOI: 10.1128/MMBR.05021-11
Diversity in genetic in vivo methods for protein-protein interaction studies: from the yeast two-hybrid system to the mammalian split-luciferase system
Abstract
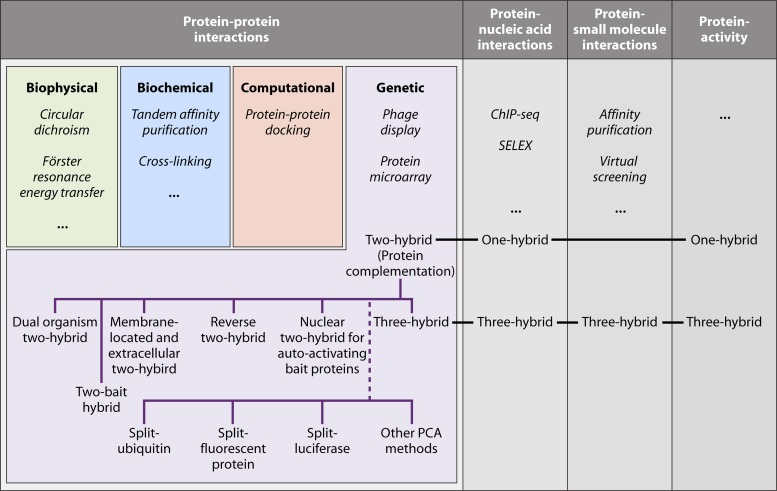
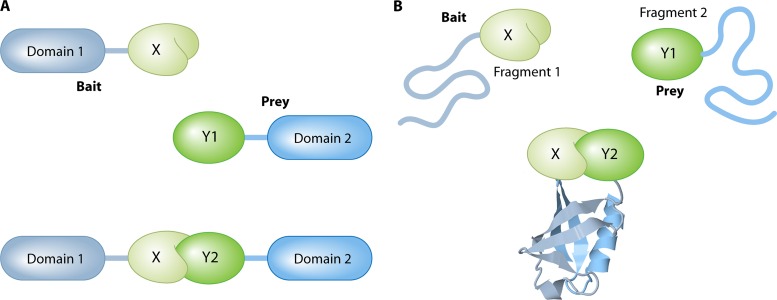
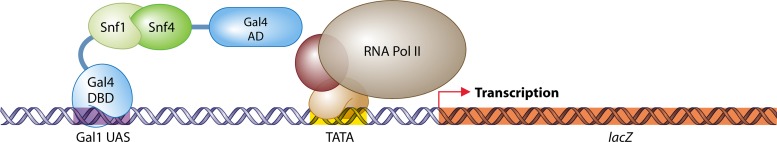
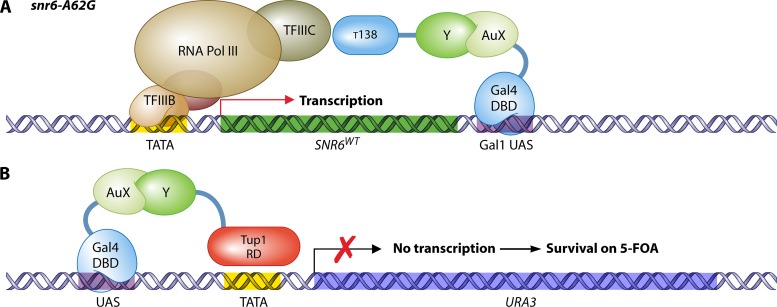
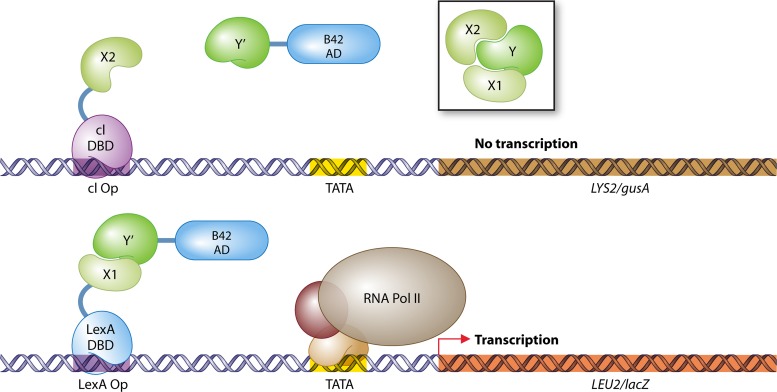
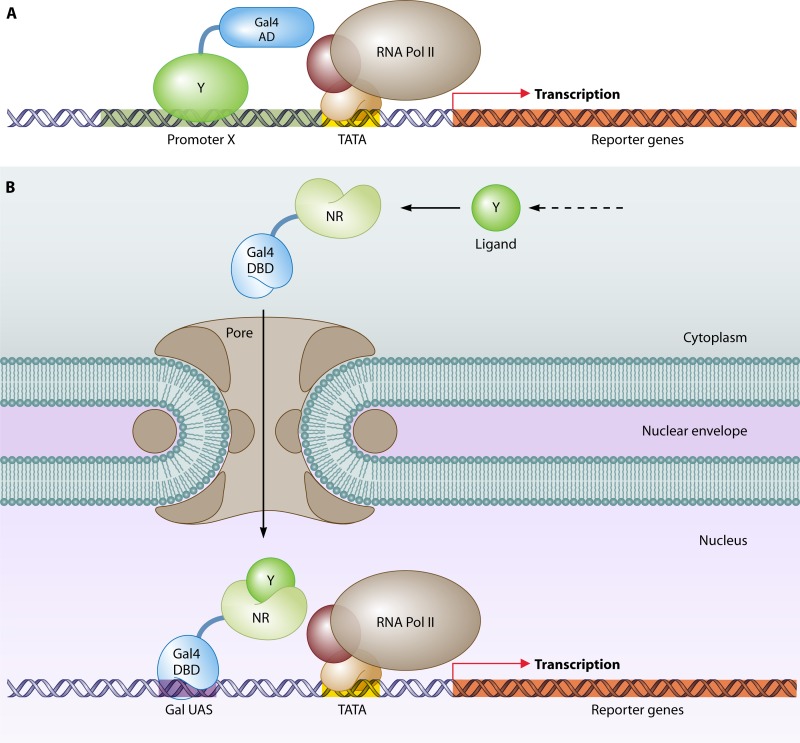
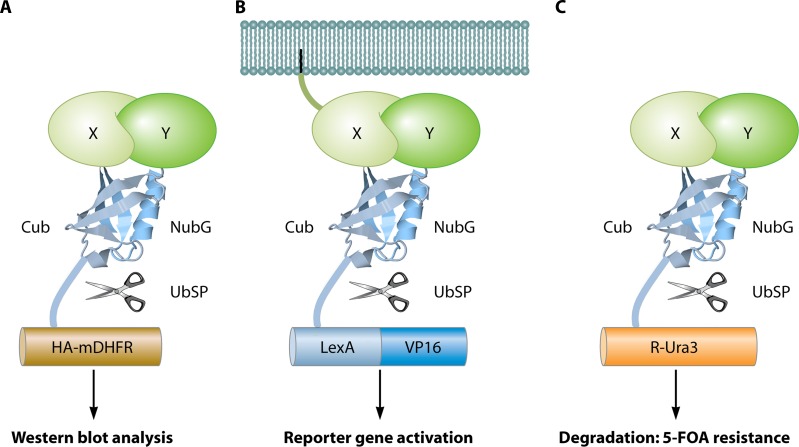
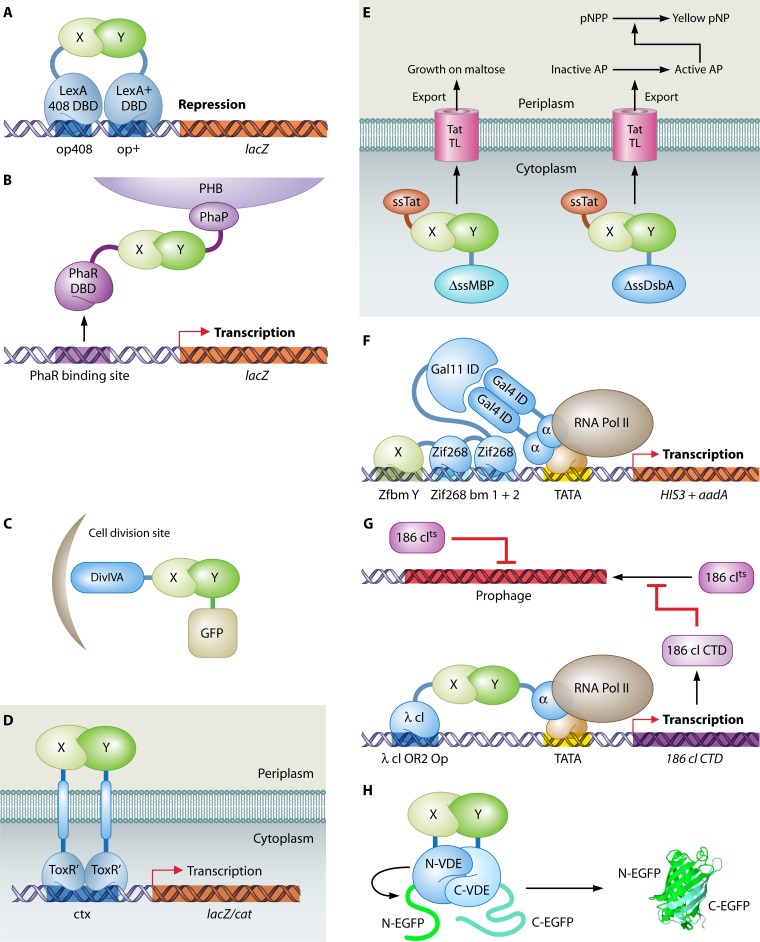
The yeast two-hybrid system pioneered the field of in vivo protein-protein interaction methods and undisputedly gave rise to a palette of ingenious techniques that are constantly pushing further the limits of the original method. Sensitivity and selectivity have improved because of various technical tricks and experimental designs. Here we present an exhaustive overview of the genetic approaches available to study in vivo binary protein interactions, based on two-hybrid and protein fragment complementation assays. These methods have been engineered and employed successfully in microorganisms such as Saccharomyces cerevisiae and Escherichia coli, but also in higher eukaryotes. From single binary pairwise interactions to whole-genome interactome mapping, the self-reassembly concept has been employed widely. Innovative studies report the use of proteins such as ubiquitin, dihydrofolate reductase, and adenylate cyclase as reconstituted reporters. Protein fragment complementation assays have extended the possibilities in protein-protein interaction studies, with technologies that enable spatial and temporal analyses of protein complexes. In addition, one-hybrid and three-hybrid systems have broadened the types of interactions that can be studied and the findings that can be obtained. Applications of these technologies are discussed, together with the advantages and limitations of the available assays.
Figures
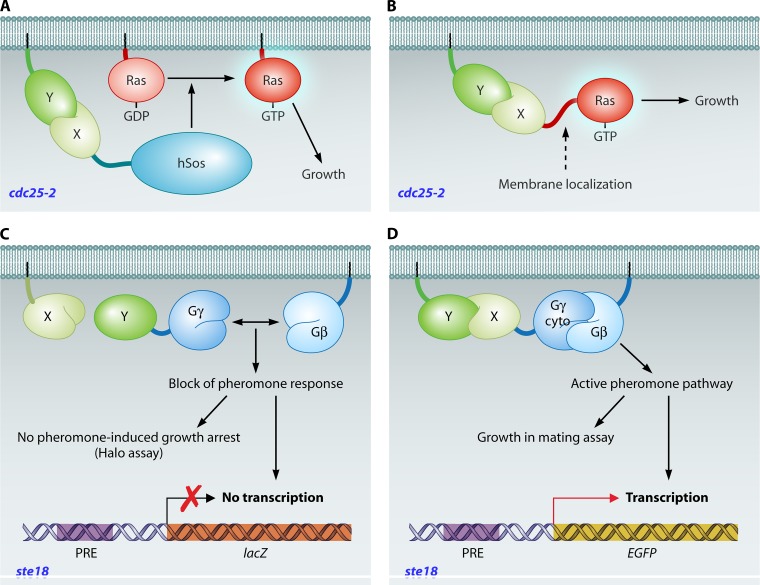
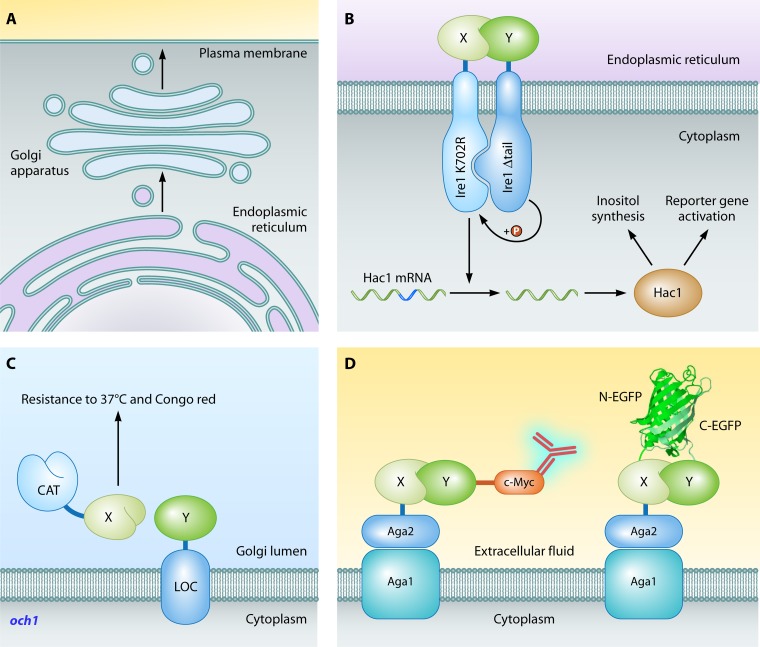
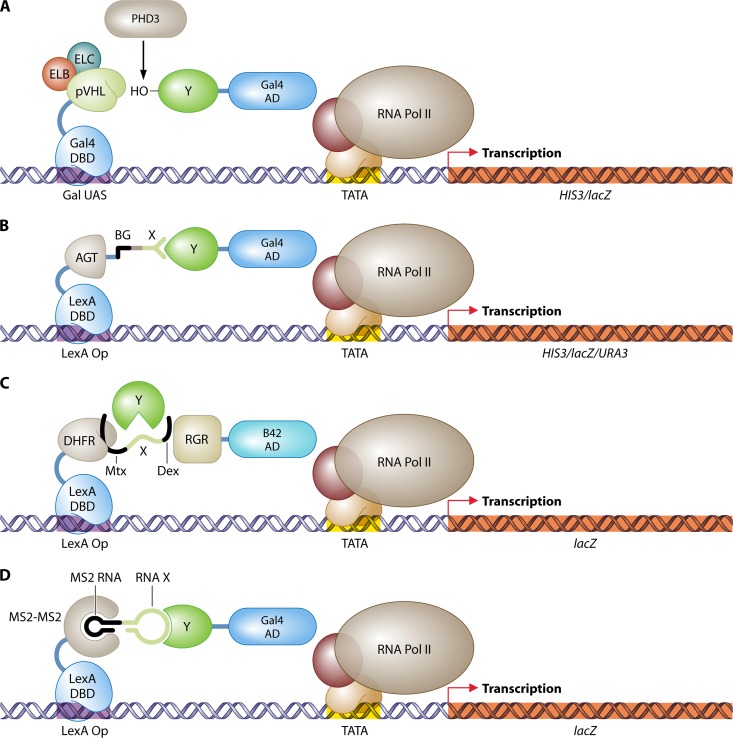
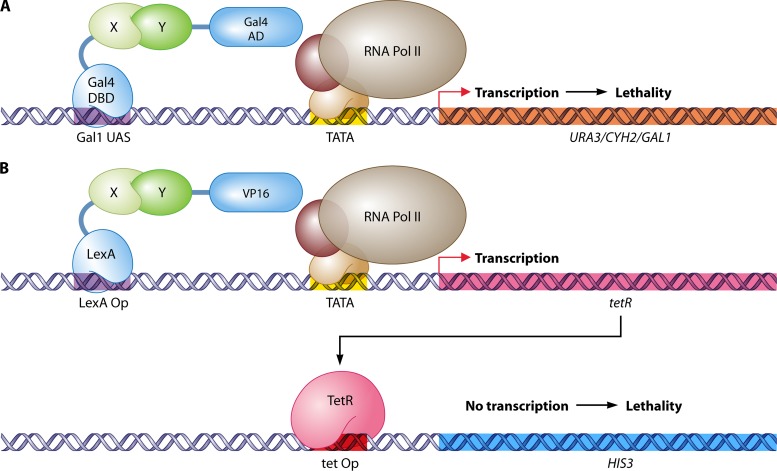
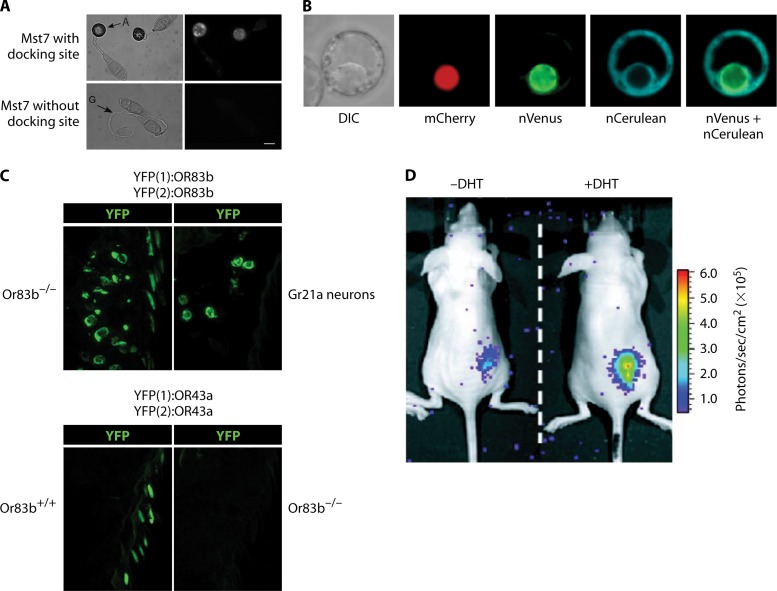
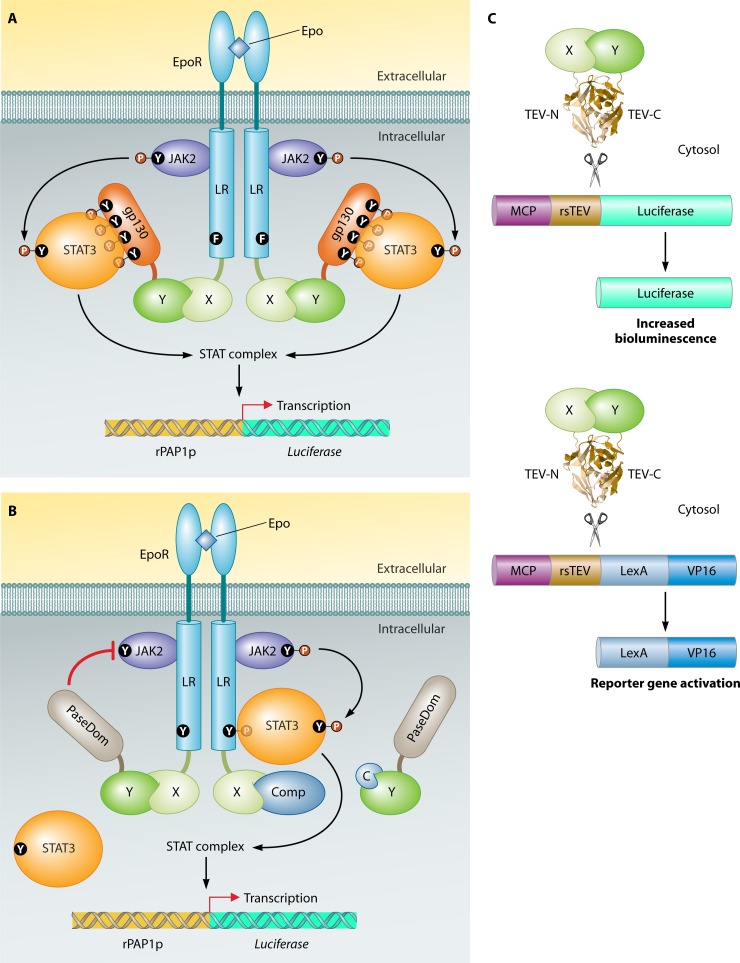




References
-
- Abida WM, Carter BT, Althoff EA, Lin H, Cornish VW. 2002. Receptor-dependence of the transcription read-out in a small-molecule three-hybrid system. Chembiochem 3:887–895 - PubMed
-
- Aguilera C, et al. 2011. c-Jun N-terminal phosphorylation antagonises recruitment of the Mbd3/NuRD repressor complex. Nature 469:231–235 - PubMed
-
- Aho S, Arfman A, Pummi T, Uitto J. 1997. A novel reporter gene MEL1 for the yeast two-hybrid system. Anal. Biochem. 253:270–272 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
