

Review
doi: 10.1083/jcb.201111146.
Scoring a backstage pass: mechanisms of ciliogenesis and ciliary access
Affiliations
- PMID: 22689651
- PMCID: PMC3373398
- DOI: 10.1083/jcb.201111146
Item in Clipboard
Review
Scoring a backstage pass: mechanisms of ciliogenesis and ciliary access
J Cell Biol.
.
Abstract
Cilia are conserved, microtubule-based cell surface projections that emanate from basal bodies, membrane-docked centrioles. The beating of motile cilia and flagella enables cells to swim and epithelia to displace fluids. In contrast, most primary cilia do not beat but instead detect environmental or intercellular stimuli. Inborn defects in both kinds of cilia cause human ciliopathies, diseases with diverse manifestations such as heterotaxia and kidney cysts. These diseases are caused by defects in ciliogenesis or ciliary function. The signaling functions of cilia require regulation of ciliary composition, which depends on the control of protein traffic into and out of cilia.
Figures
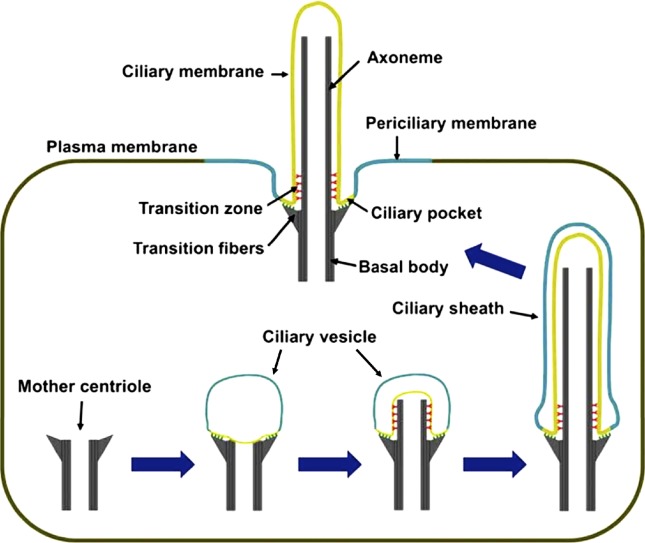
Ciliogenesis. Cilium formation starts when a mother centriole contacts a ciliary vesicle. Axonemes elongate at their tips and so are constructed from proximal to distal, with the most proximal region giving rise to the transition zone. The ciliary vesicle grows with the axoneme and gives rise to the ciliary sheath, whose fusion with the plasma membrane externalizes the cilium and transforms the outer sheath into the periciliary membrane.
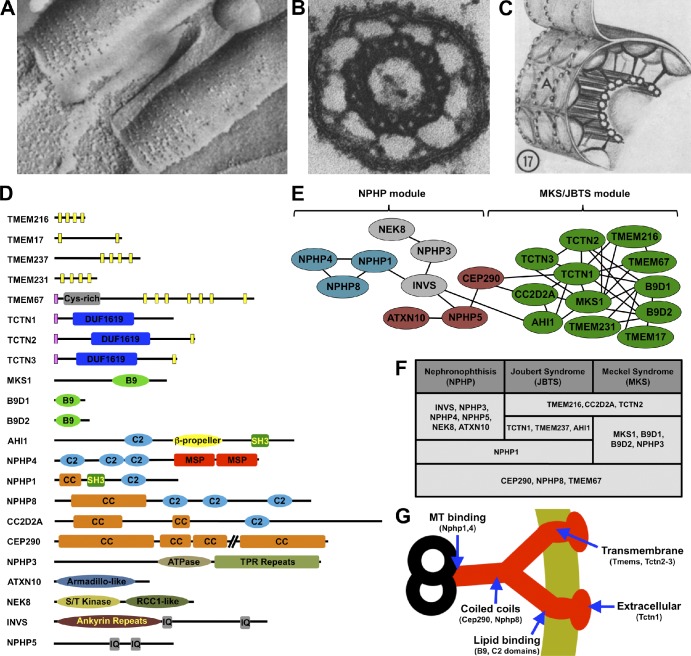
The transition zone. (A) Freeze-etch electron micrograph of tracheal epithelial cilia. The ciliary necklaces are at the ciliary base. (B) Electron micrograph of a cross section through a transition zone of a mollusk gill cilium, showing the microtubule doublets connected to the ciliary membrane by nine Y links. (C) Diagram of the transition zone, showing the Y links connecting the microtubules to the ciliary necklace. The A indicates the convex freeze-fracture face of the membrane. A–C are obtained from Gilula and Satir (1972). (D) Domain structure of transition zone components. The Tectonic proteins (TCTN1–3) share a signal peptide (pink bars) and a cysteine-rich Tectonic domain. Transition zone transmembrane proteins (TMEMs) include TCTN2, TCTN3, TMEM17, TMEM67, TMEM231, and TMEM237 (predicted transmembrane helices are shown as yellow bars). MKS1, B9D1, and B9D2 share B9 domains related to lipid-binding C2 domains. Several transition zone proteins contain C2 domains, coiled-coil (CC) domains, or both. Inversin (INVS) and NPHP5 have calmodulin-binding IQ motifs. NPHP4 contains two major sperm protein (MSP) domains, NPHP3 is an ATPase with tetratricopeptide repeat (TPR) domains, NEK8 is a serine/threonine kinase, and ATXN10 contains Armadillo repeats. (E) The transition zone protein interaction network. Genetic experiments in C. elegans reveal two main functional modules in this network. The first module is mostly comprised of genes, the human homologues of which are implicated in NPHP, whereas the second contains genes associated with MKS and JBTS. These functional modules closely match the results of biochemical experiments in mammalian cells. According to these, proteins in the MKS–JBTS module (green) mostly interact with other proteins within the same module and only rarely with those in the NPHP module (each line designates a reported protein–protein interaction). The NPHP module consists of several interconnected complexes, shown in different colors (Sang et al., 2011; van Reeuwijk et al., 2011). (F) Most known transition zone proteins are encoded by genes mutated in at least one of three related ciliopathies, NPHP, JBTS, and MKS. (G) Schematic of a Y link with a model of its composition. Nphp1 and Nphp4 bind microtubules, so they may connect Y links to microtubule doublets (Mollet et al., 2005). Cep290 and other coiled-coil proteins may form the central portion Y links. C2 and B9 domain-containing proteins are predicted to bind lipids, so they may be membrane proximal. Tctn1 also interacts with transmembrane proteins but is predicted to be on the extracellular face of the ciliary necklace, as it contains a signal peptide.
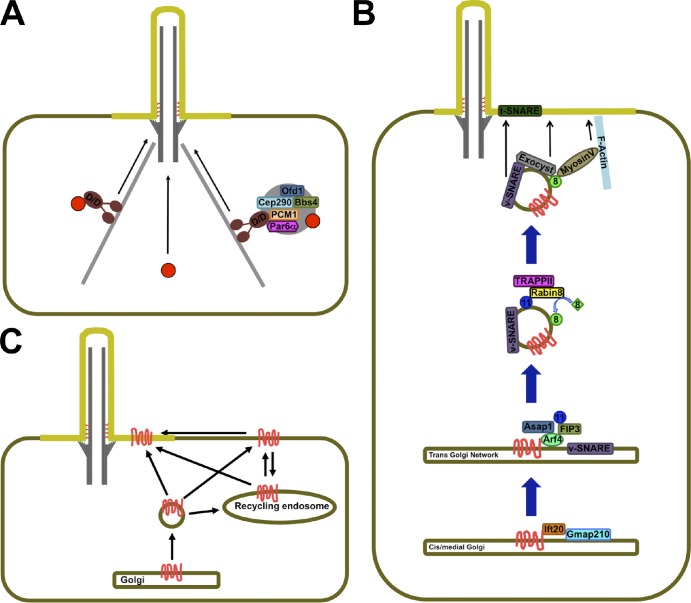
Trafficking to the ciliary base. (A) Soluble proteins (red circles) may reach the ciliary base by diffusion (center) or travel as cargo on minus end–directed microtubule motors, such as the Dynein–Dynactin complex (D/D; left). (right) Trafficking of soluble cargo may also involve centriolar satellites, large protein aggregates that may serve as assembly points, and way stations for cilium-bound proteins. (B) Trafficking of some transmembrane proteins (red) along the Golgi may be aided by IFT20, which may then hand them off to Arf4. Arf4 orchestrates the formation of cargo-containing cilium-bound vesicles that contain active Rab11 (blue), whose effector Rabin8 recruits active Rab8 (green) to the vesicle surface. Rab8 in turn recruits effectors that mediate the vesicle’s approach, tethering, and fusion with the periciliary membrane. (C) Transmembrane proteins (red) may reach the cilium laterally from the plasma membrane or aboard vesicles that fuse with the periciliary membrane. Cilia-bound vesicles may derive from the Golgi or from recycling endosomes, which themselves may receive input from the Golgi and plasma membrane.
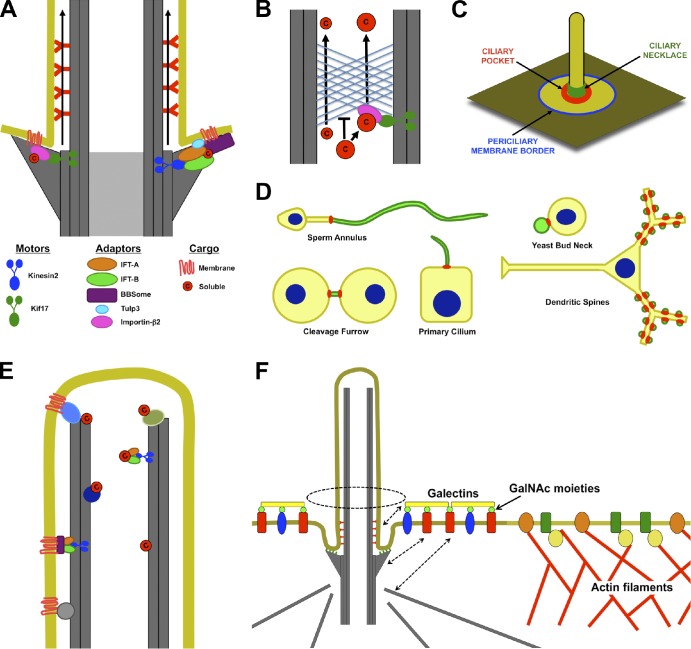
Access into and maintenance of distinct ciliary compartments. (A) Both soluble and transmembrane proteins may need to associate directly or indirectly with plus end–directed microtubule motors (Kinesin-2 or Kif17) to enter the cilium. The entrance may be between adjacent transition fibers. (B) Like the nuclear pore, the ciliary base can exclude proteins on the basis of their size. Thus, small proteins may diffuse into the cilium, but large proteins may need to associate with transport machinery to enter the cilium. (C) Potential locations for diffusion barriers preventing entrance of membrane proteins into cilia include the border between the plasma and periciliary membranes (blue), the bottom of the ciliary pocket where transition fibers anchor the basal body to the membrane (red), and the ciliary necklace (green). (D) Septin rings (red) form membrane diffusion barriers that define specific membrane compartments (green), including those of cilia, flagella, the midbody, dendritic spines, and yeast buds. (E) Soluble and membrane proteins may be retained inside cilia by directly or indirectly interacting with microtubules. (F) Selective exclusion and retention can account for differences in periciliary and plasma membrane composition. The cortical actin cytoskeleton is excluded from the region under the periciliary membrane. As a result, membrane proteins that directly or indirectly interact with actin filaments are excluded from the periciliary region. Conversely, certain proteins, such as Galectin-3, that accumulate in the periciliary region may be retained by extracytosolic interactions with the ciliary membrane or by interactions with the basal body or its associated microtubules (dashed arrows). c, cargo; GalNAc, N-acetyl-galactosamine.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
