

Conserved genetic determinant of motor organ identity in Medicago truncatula and related legumes
- PMID: 22689967
- PMCID: PMC3406810
- DOI: 10.1073/pnas.1204566109
Conserved genetic determinant of motor organ identity in Medicago truncatula and related legumes
Abstract
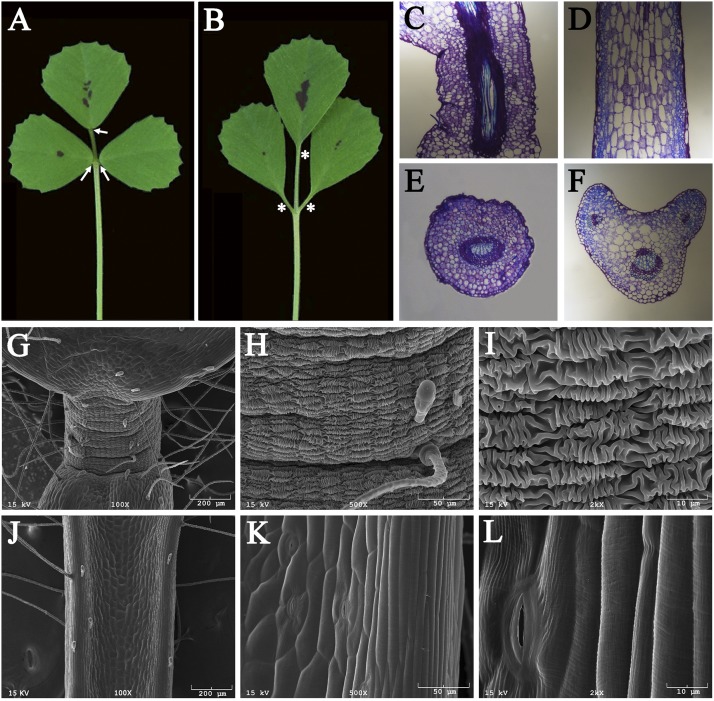
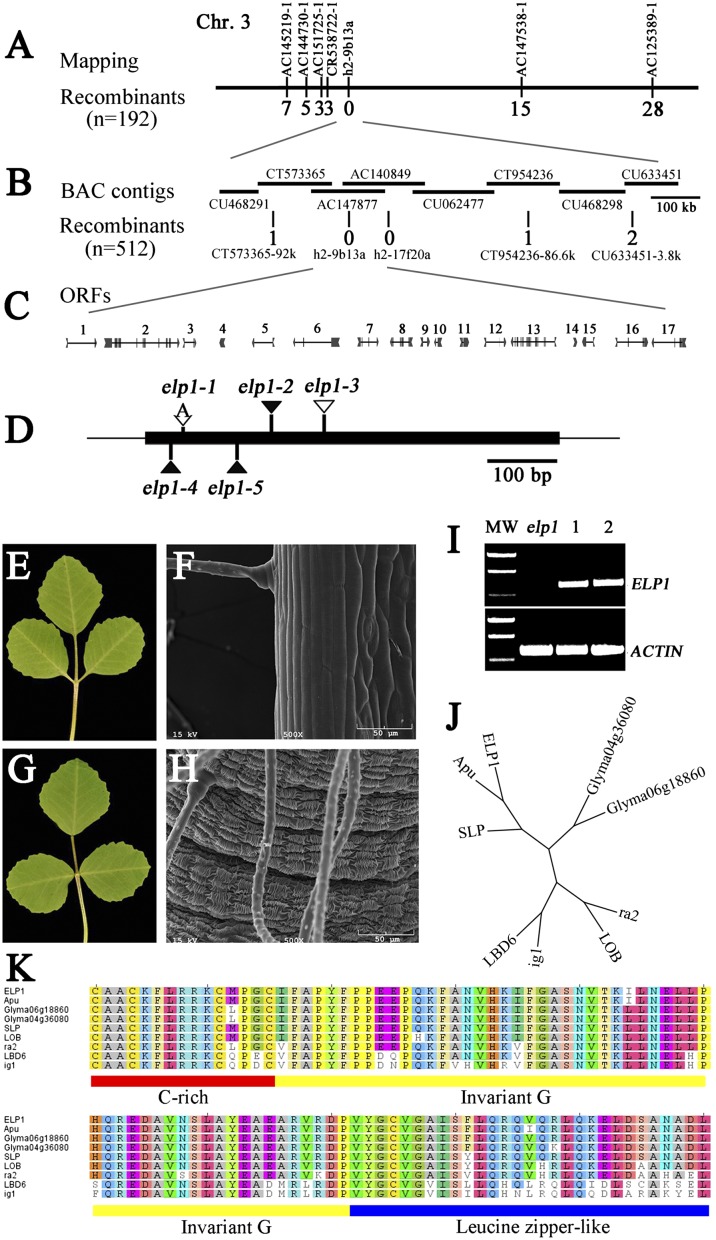
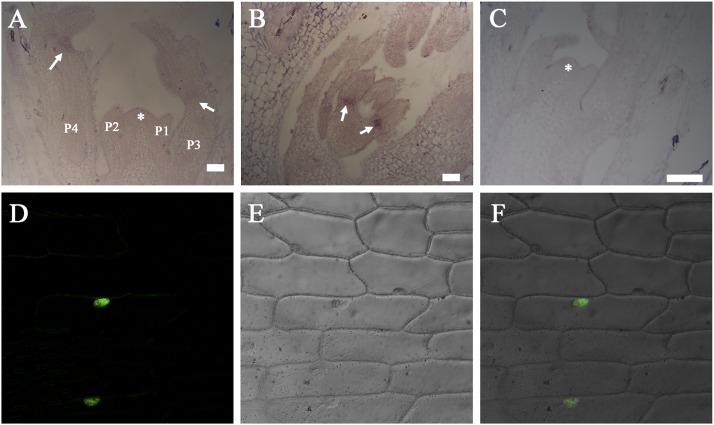
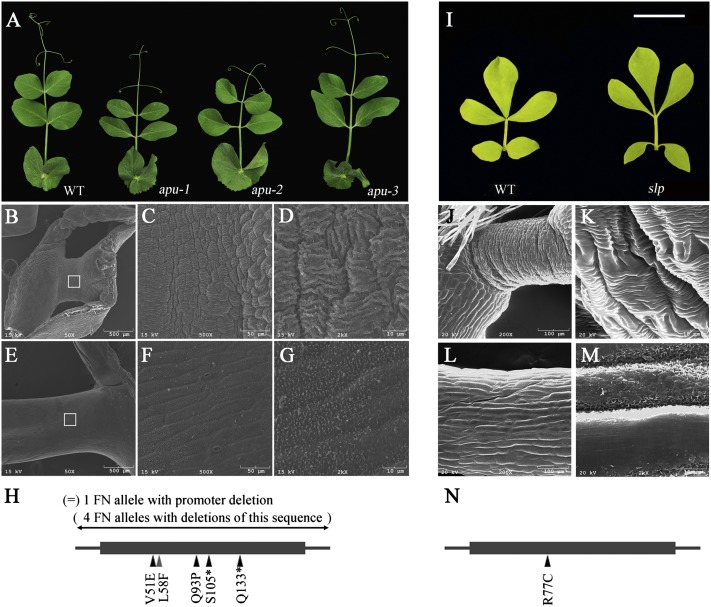
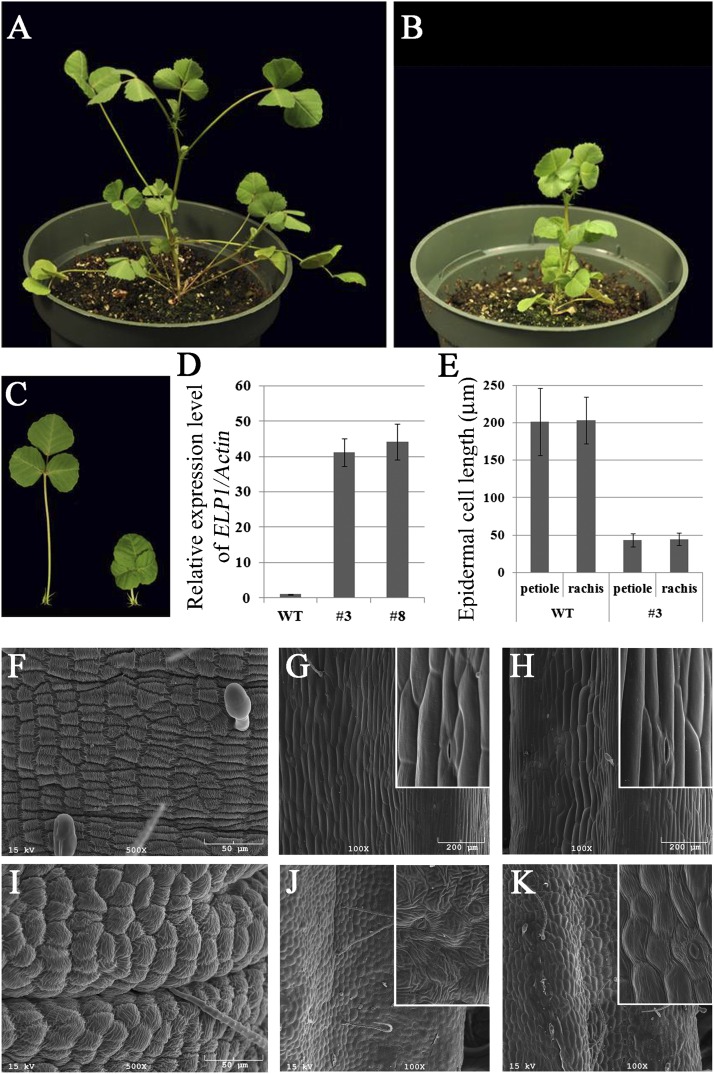
Plants exhibit various kinds of movements that have fascinated scientists and the public for centuries. Physiological studies in plants with the so-called motor organ or pulvinus suggest that cells at opposite sides of the pulvinus mediate leaf or leaflet movements by swelling and shrinking. How motor organ identity is determined is unknown. Using a genetic approach, we isolated a mutant designated elongated petiolule1 (elp1) from Medicago truncatula that fails to fold its leaflets in the dark due to loss of motor organs. Map-based cloning indicated that ELP1 encodes a putative plant-specific LOB domain transcription factor. RNA in situ analysis revealed that ELP1 is expressed in primordial cells that give rise to the motor organ. Ectopic expression of ELP1 resulted in dwarf plants with petioles and rachises reduced in length, and the epidermal cells gained characteristics of motor organ epidermal cells. By identifying ELP1 orthologs from other legume species, namely pea (Pisum sativum) and Lotus japonicus, we show that this motor organ identity is regulated by a conserved molecular mechanism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Genetic basis of the "sleeping leaves" revealed.Proc Natl Acad Sci U S A. 2012 Jul 17;109(29):11474-5. doi: 10.1073/pnas.1209532109. Epub 2012 Jul 6. Proc Natl Acad Sci U S A. 2012. PMID: 22773806 Free PMC article. No abstract available.
References
-
- Koller D. The Restless Plant. Cambridge, MA: Harvard Univ Press; 2011.
-
- Darwin C. The Power of Movement in Plants. New York: D. Appleton and Company; 1897.
-
- Hangarter R. Plants-In-Motion. 2000. Available at http://plantsinmotion.bio.indiana.edu/plantmotion/starthere.html.
-
- Chen R, Guan C, Boonsirichai K, Masson PH. Complex physiological and molecular processes underlying root gravitropism. Plant Mol Biol. 2002;49:305–317. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
