

Autocrine/paracrine TGF-β1 inhibits Langerhans cell migration
- PMID: 22689996
- PMCID: PMC3387113
- DOI: 10.1073/pnas.1119178109
Autocrine/paracrine TGF-β1 inhibits Langerhans cell migration
Abstract
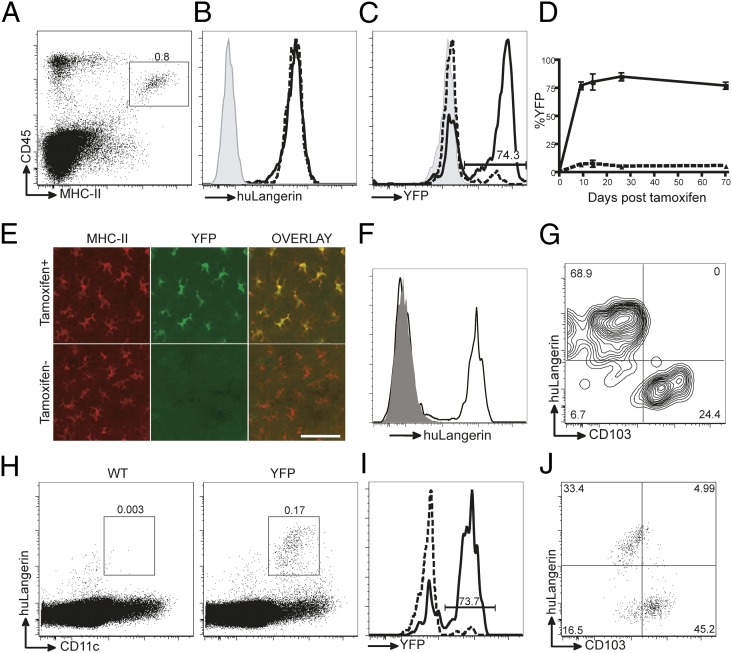
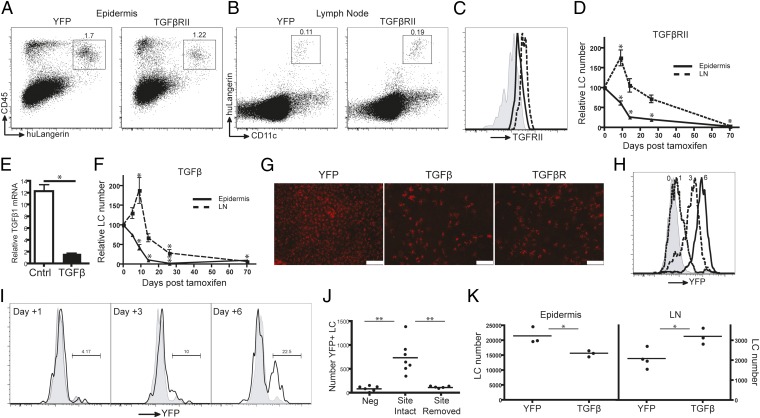
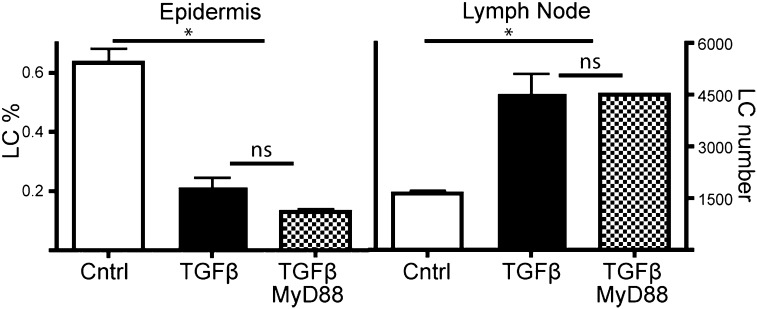
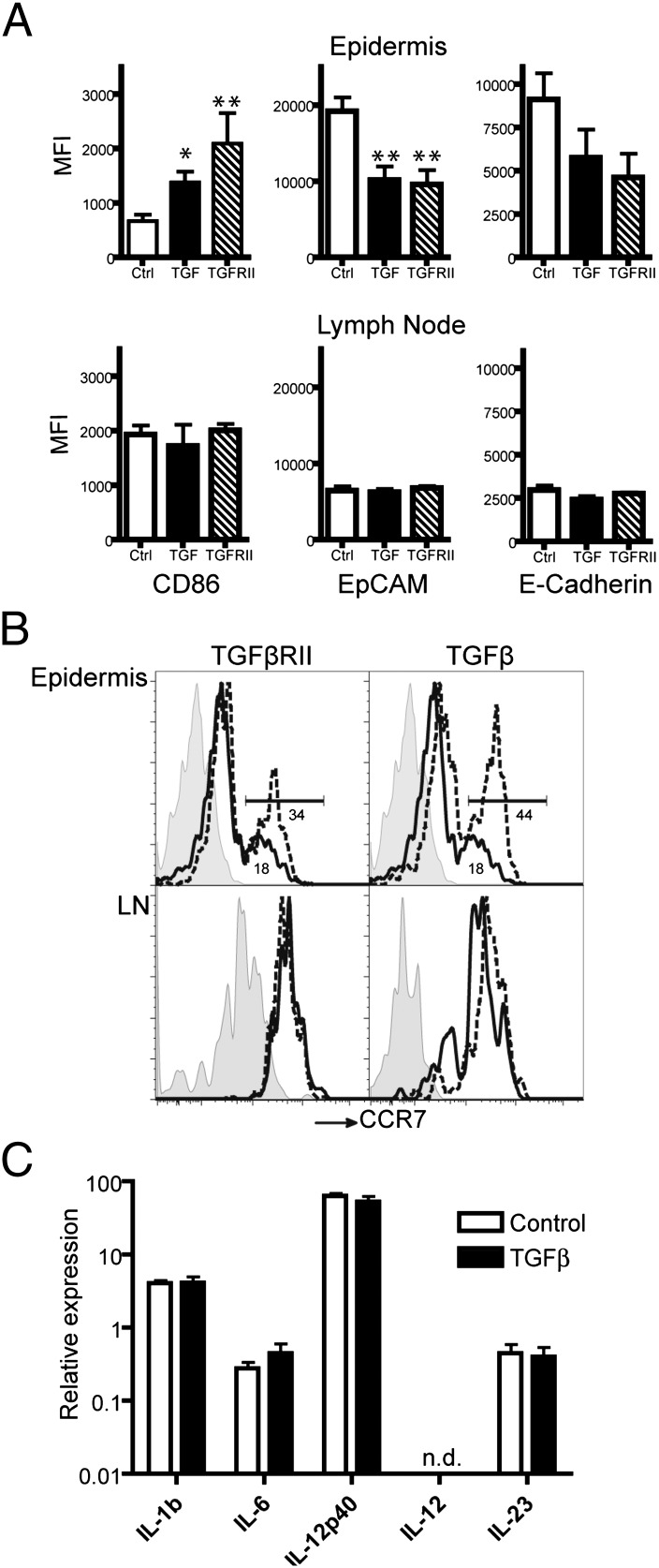
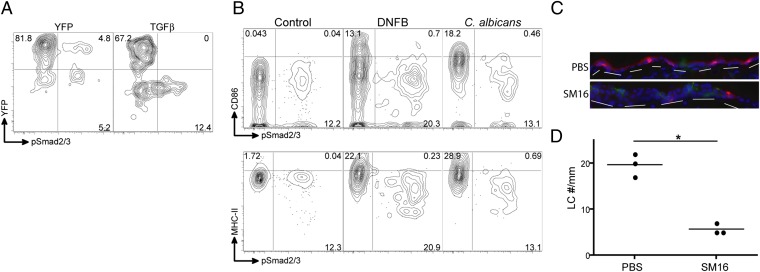
Langerhans cells (LCs) are skin-resident dendritic cells (DC) located in the epidermis that migrate to skin-draining lymph nodes during the steady state and in response to inflammatory stimuli. TGF-β1 is a critical immune regulator that is highly expressed by LCs. The ability to test the functional importance of LC-derived TGF-β1 is complicated by the requirement of TGF-β1 for LC development and by the absence of LCs in mice with an LC-specific ablation of TGF-β1 or its receptor. To overcome these problems, we have engineered transgenic huLangerin-CreER(T2) mice that allow for inducible LC-specific excision. Highly efficient and LC-specific expression was confirmed in mice bred onto a YFP Cre reporter strain. We next generated huLangerin-CreER(T2) × TGF-βRII(fl) and huLangerin-CreER(T2) × TGF-β1(fl) mice. Excision of the TGFβRII or TGFβ1 genes induced mass migration of LCs to the regional lymph node. Expression of costimulatory markers and inflammatory cytokines was unaffected, consistent with homeostatic migration. In addition, levels of p-SMAD2/3 were decreased in LCs from wild-type mice before inflammation-induced migration. We conclude that TGF-β1 acts directly on LCs in an autocrine/paracrine manner to inhibit steady-state and inflammation-induced migration. This is a readily targetable pathway with potential therapeutic implications for skin disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hemmi H, et al. Skin antigens in the steady state are trafficked to regional lymph nodes by transforming growth factor-beta1-dependent cells. Int Immunol. 2001;13:695–704. - PubMed
-
- Merad M, Ginhoux F, Collin M. Origin, homeostasis and function of Langerhans cells and other langerin-expressing dendritic cells. Nat Rev Immunol. 2008;8:935–947. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
