

Convergent Evolution of Calcineurin Pathway Roles in Thermotolerance and Virulence in Candida glabrata
- PMID: 22690377
- PMCID: PMC3362297
- DOI: 10.1534/g3.112.002279
Convergent Evolution of Calcineurin Pathway Roles in Thermotolerance and Virulence in Candida glabrata
Erratum in
-
Correction to: Convergent Evolution of Calcineurin Pathway Roles in Thermotolerance and Virulence in Candida glabrata.G3 (Bethesda). 2025 Aug 6;15(8):jkaf124. doi: 10.1093/g3journal/jkaf124. G3 (Bethesda). 2025. PMID: 40489317 Free PMC article. No abstract available.
Abstract
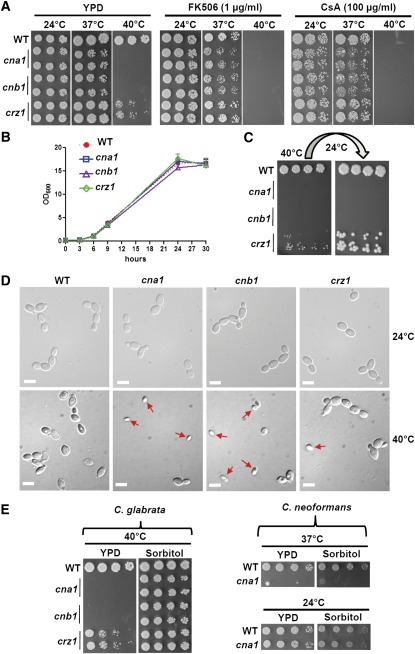
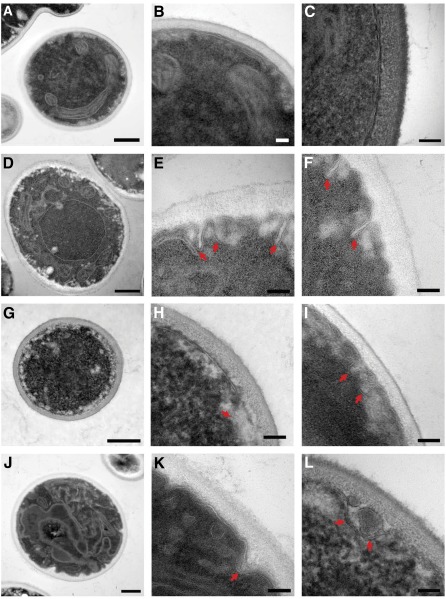
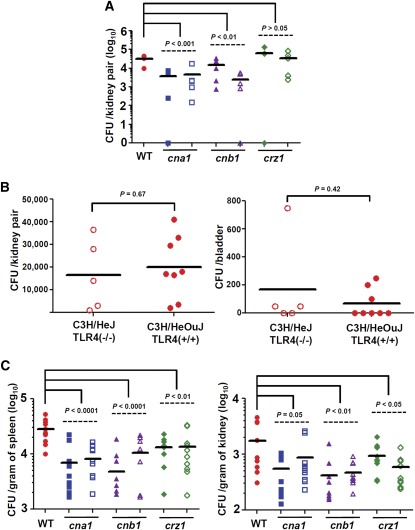
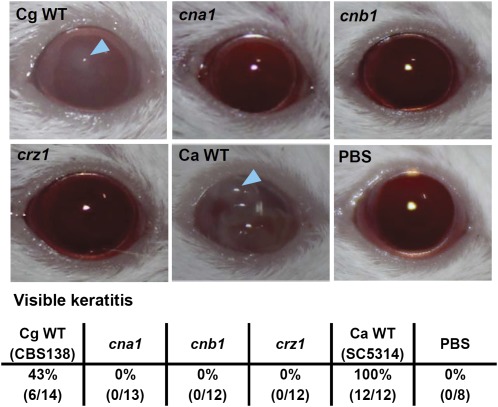
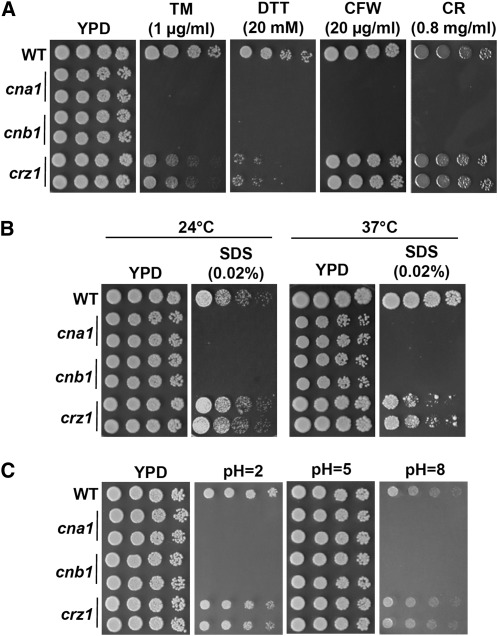
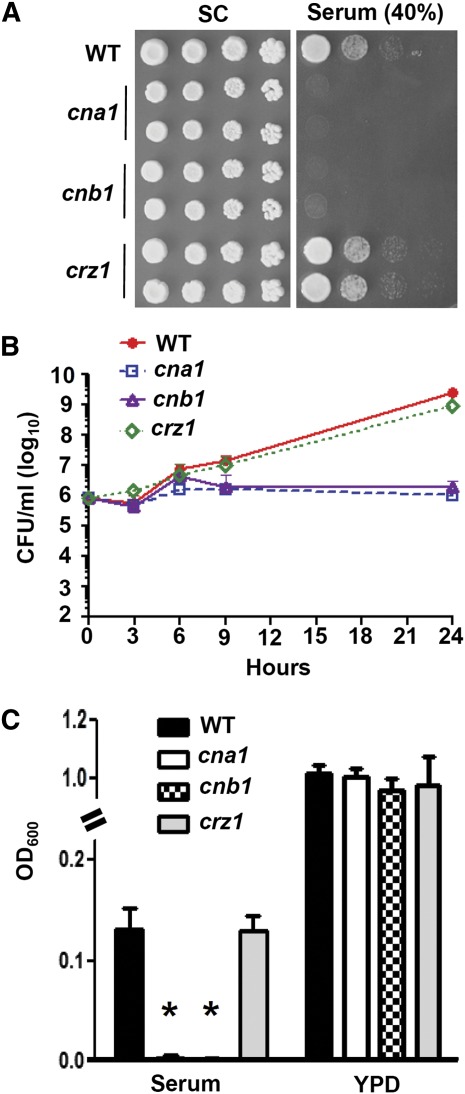
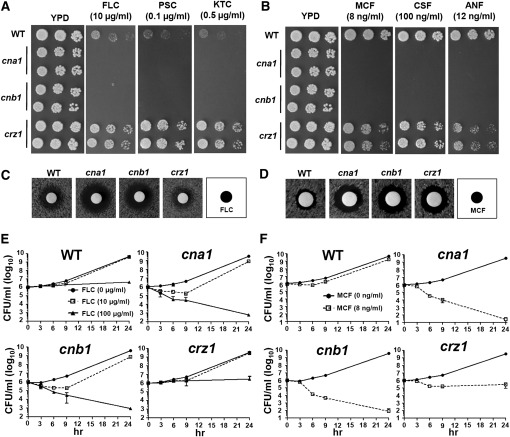
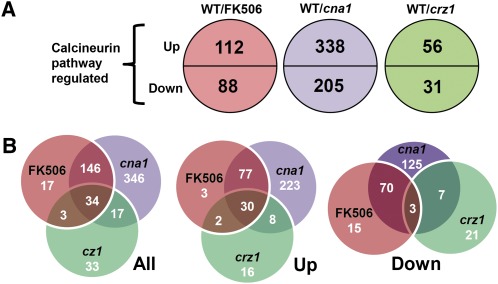
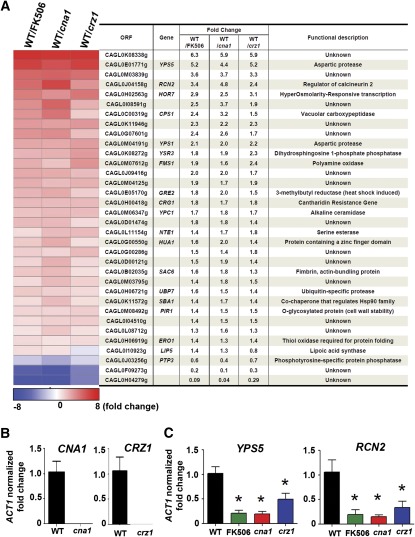
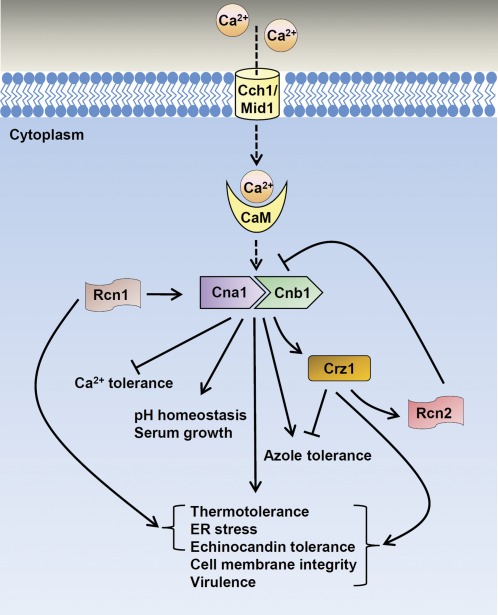
Candida glabrata is an emerging human fungal pathogen that is frequently drug tolerant, resulting in difficulties in treatment and a higher mortality in immunocompromised patients. The calcium-activated protein phosphatase calcineurin plays critical roles in controlling drug tolerance, hyphal growth, and virulence in diverse fungal pathogens via distinct mechanisms involving survival in serum or growth at host temperature (37° and higher). Here, we comprehensively studied the calcineurin signaling cascade in C. glabrata and found novel and uncharacterized functions of calcineurin and its downstream target Crz1 in governing thermotolerance, intracellular architecture, and pathogenesis in murine ocular, urinary tract, and systemic infections. This represents a second independent origin of a role for calcineurin in thermotolerant growth of a major human fungal pathogen, distinct from that which arose independently in Cryptococcus neoformans. Calcineurin also promotes survival of C. glabrata in serum via mechanisms distinct from C. albicans and thereby enables establishment of tissue colonization in a murine systemic infection model. To understand calcineurin signaling in detail, we performed global transcript profiling analysis and identified calcineurin- and Crz1-dependent genes in C. glabrata involved in cell wall biosynthesis, heat shock responses, and calcineurin function. Regulators of calcineurin (RCN) are a novel family of calcineurin modifiers, and two members of this family were identified in C. glabrata: Rcn1 and Rcn2. Our studies demonstrate that Rcn2 expression is controlled by calcineurin and Crz1 to function as a feedback inhibitor of calcineurin in a circuit required for calcium tolerance in C. glabrata. In contrast, the calcineurin regulator Rcn1 activates calcineurin signaling. Interestingly, neither Rcn1 nor Rcn2 is required for virulence in a murine systemic infection model. Taken together, our findings show that calcineurin signaling plays critical roles in thermotolerance and virulence, and that Rcn1 and Rcn2 have opposing functions in controlling calcineurin signaling in C. glabrata.
Keywords: Crz1; ER stress; Rcn1; Rcn2; calcium; calmodulin; cell wall integrity; drug tolerance; ocular infection; pH homeostasis; phosphatase; thermotolerance; urinary tract infection; virulence.
Figures

Similar articles
-
The calcineurin pathway regulates extreme thermotolerance, cell membrane and wall integrity, antifungal resistance, and virulence in Candida auris.PLoS Pathog. 2025 Jul 28;21(7):e1013363. doi: 10.1371/journal.ppat.1013363. eCollection 2025 Jul. PLoS Pathog. 2025. PMID: 40720551 Free PMC article.
-
Calcineurin signaling: lessons from Candida species.FEMS Yeast Res. 2015 Jun;15(4):fov016. doi: 10.1093/femsyr/fov016. Epub 2015 Apr 15. FEMS Yeast Res. 2015. PMID: 25878052 Review.
-
Calcineurin controls drug tolerance, hyphal growth, and virulence in Candida dubliniensis.Eukaryot Cell. 2011 Jun;10(6):803-19. doi: 10.1128/EC.00310-10. Epub 2011 Apr 29. Eukaryot Cell. 2011. PMID: 21531874 Free PMC article.
-
Calcineurin controls hyphal growth, virulence, and drug tolerance of Candida tropicalis.Eukaryot Cell. 2014 Jul;13(7):844-54. doi: 10.1128/EC.00302-13. Epub 2014 Jan 17. Eukaryot Cell. 2014. PMID: 24442892 Free PMC article.
-
CRZ1 transcription factor is involved in cell survival, stress tolerance, and virulence in fungi.J Biosci. 2022;47:66. J Biosci. 2022. PMID: 36408540 Review.
Cited by
-
Sesamol: a natural phenolic compound with promising anticandidal potential.J Pathog. 2014;2014:895193. doi: 10.1155/2014/895193. Epub 2014 Dec 9. J Pathog. 2014. PMID: 25574401 Free PMC article.
-
Genetic analysis of Hsp90 function in Cryptococcus neoformans highlights key roles in stress tolerance and virulence.Genetics. 2022 Jan 4;220(1):iyab164. doi: 10.1093/genetics/iyab164. Genetics. 2022. PMID: 34849848 Free PMC article.
-
Of mice, flies--and men? Comparing fungal infection models for large-scale screening efforts.Dis Model Mech. 2015 May;8(5):473-86. doi: 10.1242/dmm.019901. Epub 2015 Mar 18. Dis Model Mech. 2015. PMID: 25786415 Free PMC article.
-
The Unfolded Protein Response Pathway in the Yeast Kluyveromyces lactis. A Comparative View among Yeast Species.Cells. 2018 Aug 14;7(8):106. doi: 10.3390/cells7080106. Cells. 2018. PMID: 30110882 Free PMC article. Review.
-
Calcineurin plays key roles in the dimorphic transition and virulence of the human pathogenic zygomycete Mucor circinelloides.PLoS Pathog. 2013;9(9):e1003625. doi: 10.1371/journal.ppat.1003625. Epub 2013 Sep 5. PLoS Pathog. 2013. PMID: 24039585 Free PMC article.
References
-
- Adams A. E., Pringle J. R., 1991. Staining of actin with fluorochrome-conjugated phalloidin. Methods Enzymol. 194: 729–731 - PubMed
-
- Al-Assiri A., Al-Jastaneiah S., Al-Khalaf A., Al-Fraikh H., Wagoner M. D., 2006. Late-onset donor-to-host transmission of Candida glabrata following corneal transplantation. Cornea 25: 123–125 - PubMed
-
- Bairwa G., Kaur R., 2011. A novel role for a glycosylphosphatidylinositol-anchored aspartyl protease, CgYps1, in the regulation of pH homeostasis in Candida glabrata. Mol. Microbiol. 79: 900–913 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous