

Phage-mediated acquisition of a type III secreted effector protein boosts growth of salmonella by nitrate respiration
- PMID: 22691391
- PMCID: PMC3374392
- DOI: 10.1128/mBio.00143-12
Phage-mediated acquisition of a type III secreted effector protein boosts growth of salmonella by nitrate respiration
Abstract
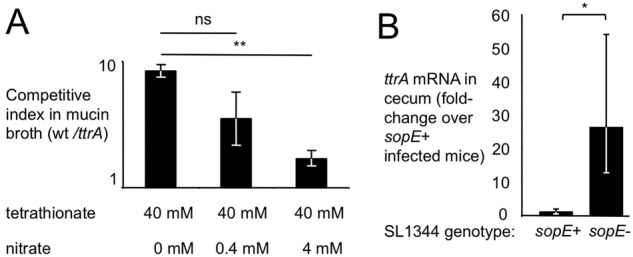
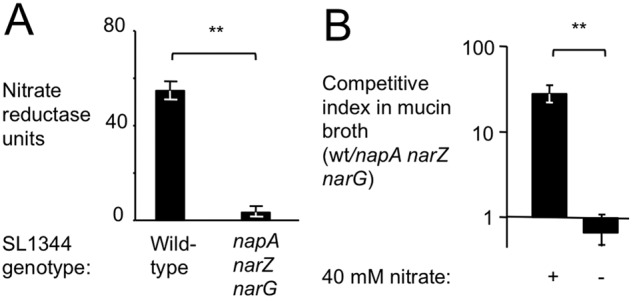
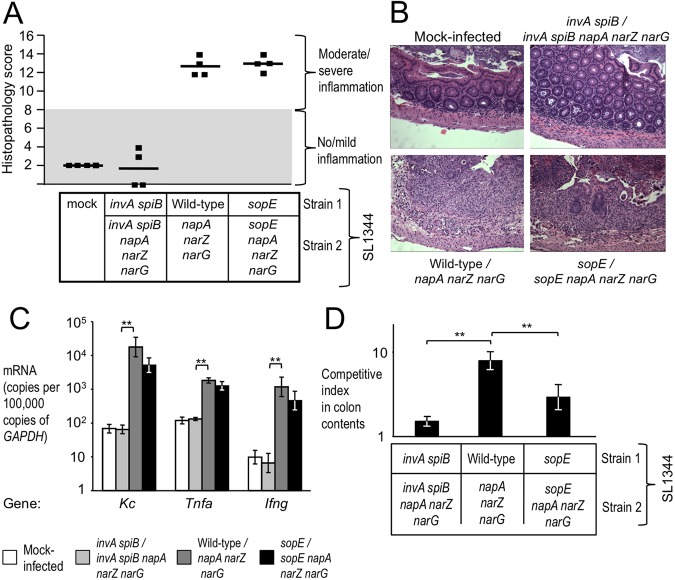
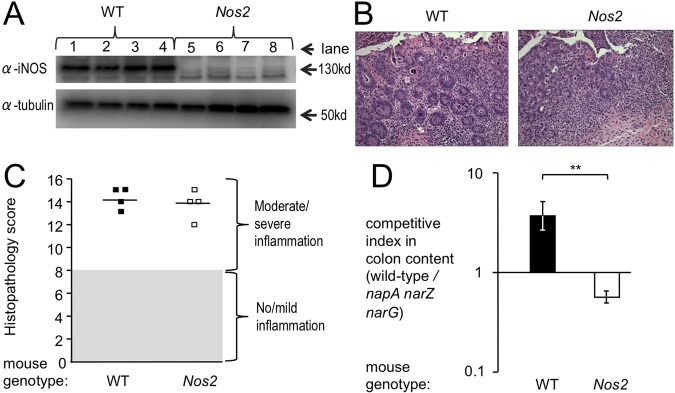
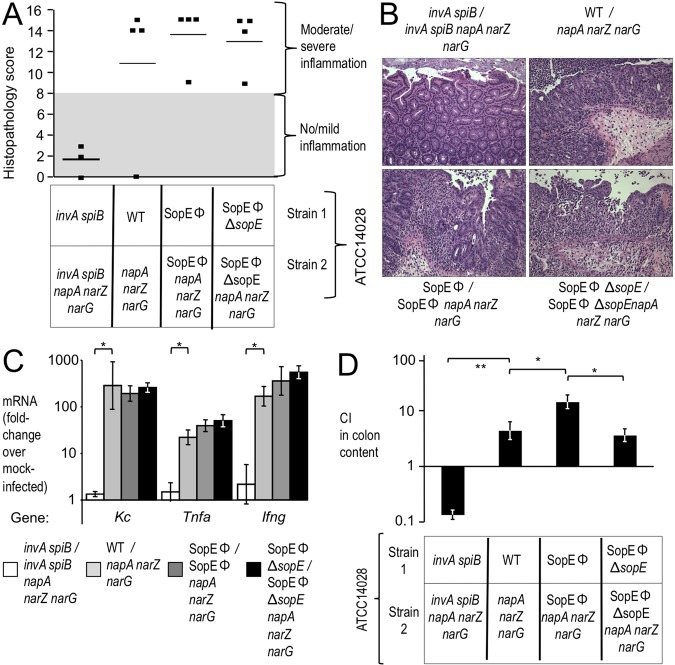
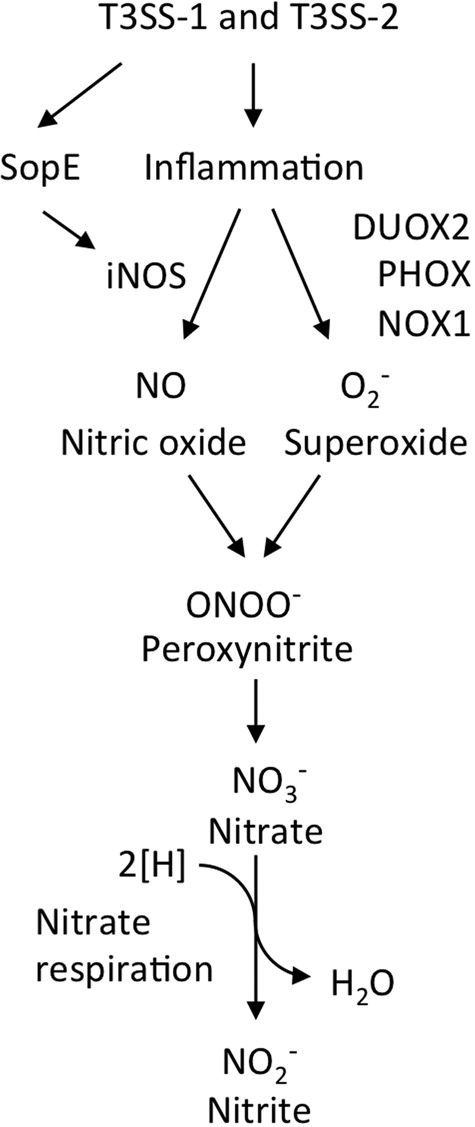
Information on how emerging pathogens can invade and persist and spread within host populations remains sparse. In the 1980s, a multidrug-resistant Salmonella enterica serotype Typhimurium clone lysogenized by a bacteriophage carrying the sopE virulence gene caused an epidemic among cattle and humans in Europe. Here we show that phage-mediated horizontal transfer of the sopE gene enhances the production of host-derived nitrate, an energetically highly valuable electron acceptor, in a mouse colitis model. In turn, nitrate fuels a bloom of S. Typhimurium in the gut lumen through anaerobic nitrate respiration while suppressing genes for the utilization of energetically inferior electron acceptors such as tetrathionate. Through this mechanism, horizontal transfer of sopE can enhance the fitness of S. Typhimurium, resulting in its significantly increased abundance in the feces. IMPORTANCE During gastroenteritis, Salmonella enterica serotype Typhimurium can use tetrathionate respiration to edge out competing microbes in the gut lumen. However, the concept that tetrathionate respiration confers a growth benefit in the inflamed gut is not broadly applicable to other host-pathogen combinations because tetrathionate respiration is a signature trait used to differentiate Salmonella serotypes from most other members of the family Enterobacteriaceae. Here we show that by acquiring the phage-carried sopE gene, S. Typhimurium can drive the host to generate an additional respiratory electron acceptor, nitrate. Nitrate suppresses genes for the utilization of energetically inferior electron acceptors such as tetrathionate while enhancing the luminal growth of S. Typhimurium through anaerobic nitrate respiration. Pathways for anaerobic nitrate respiration are widely conserved among members of the family Enterobacteriaceae, thereby making our observations relevant to other enteric pathogens whose relative abundance in the intestinal lumen increases during infection.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
