

Acetylation modulates cellular distribution and DNA sensing ability of interferon-inducible protein IFI16
- PMID: 22691496
- PMCID: PMC3387042
- DOI: 10.1073/pnas.1203447109
Acetylation modulates cellular distribution and DNA sensing ability of interferon-inducible protein IFI16
Abstract
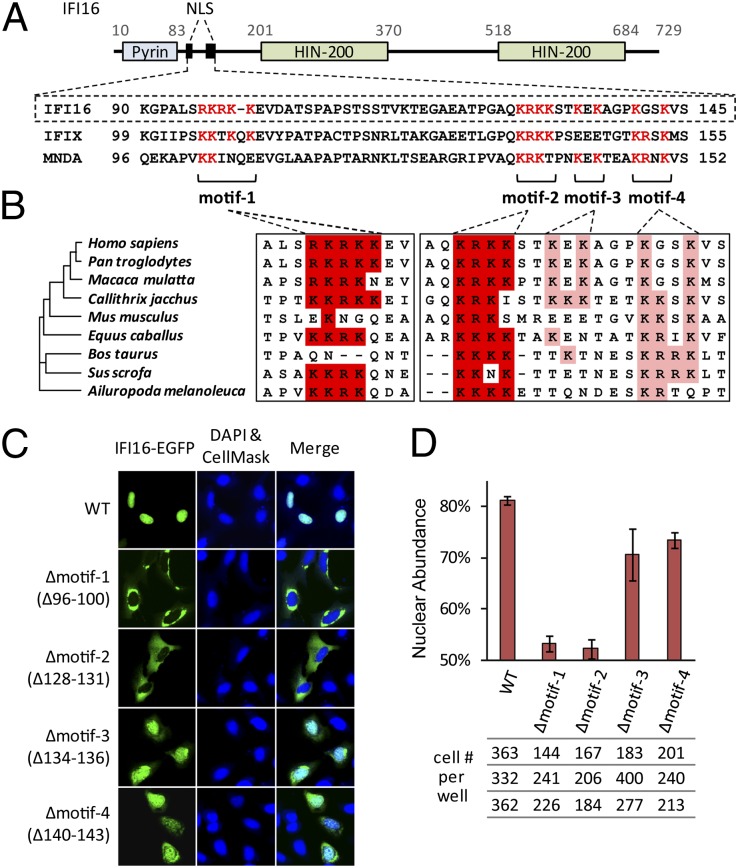
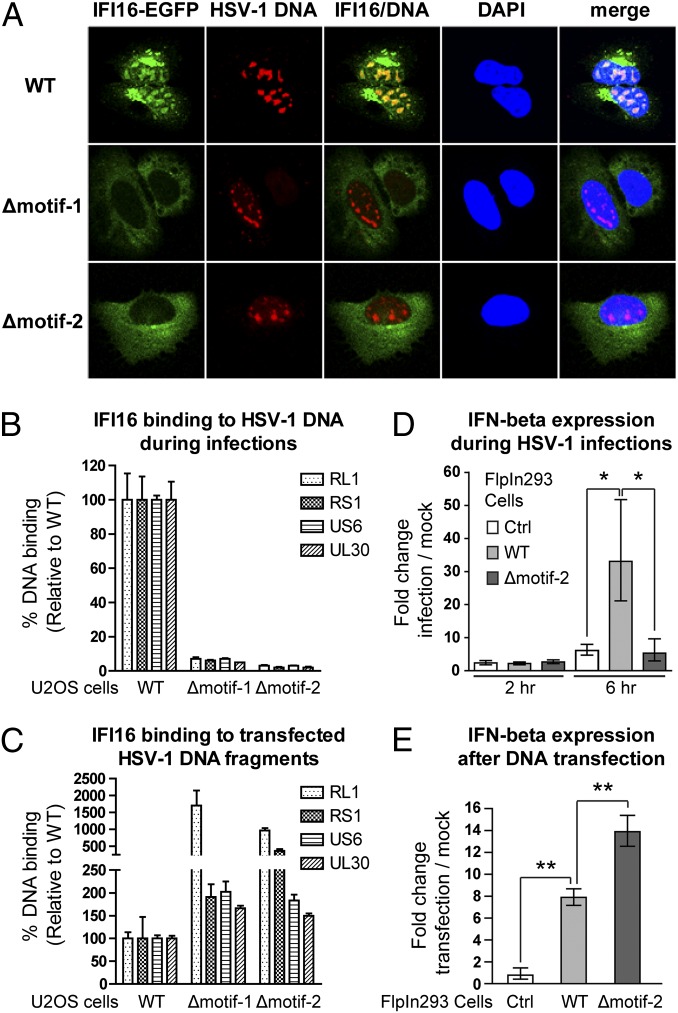
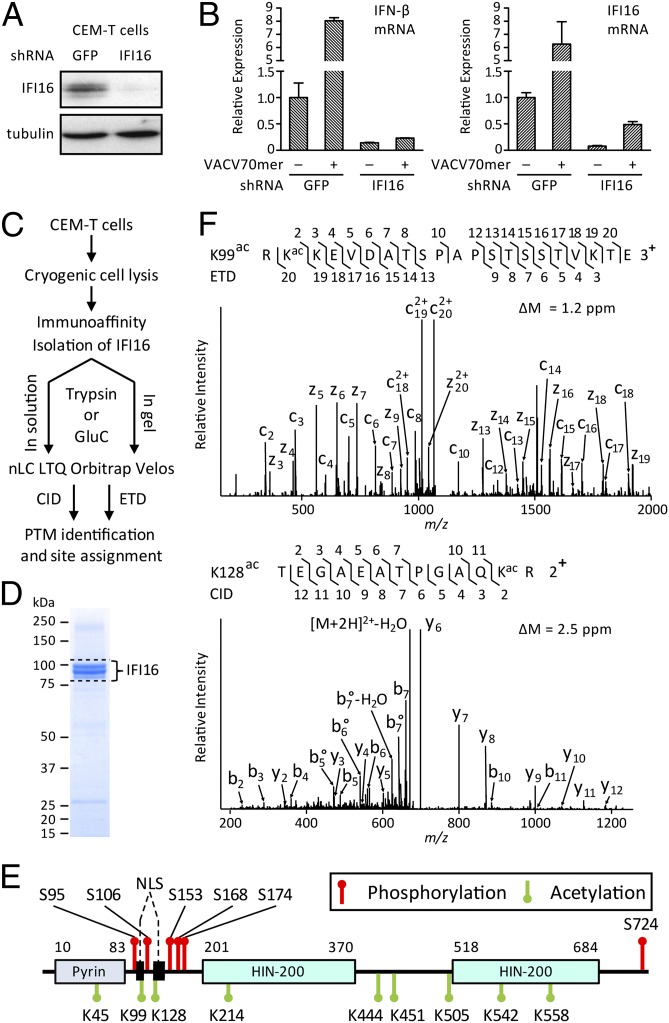
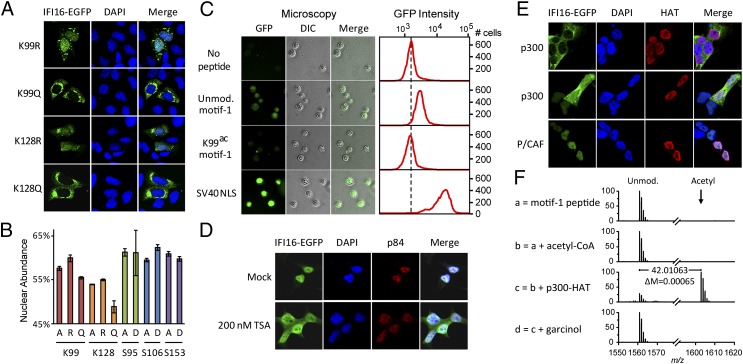
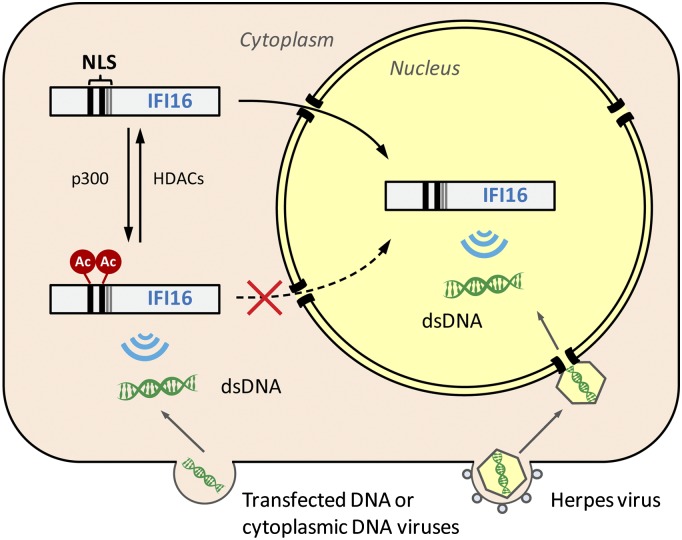
Detection of pathogenic nucleic acids is essential for mammalian innate immunity. IFN-inducible protein IFI16 has emerged as a critical sensor for detecting pathogenic DNA, stimulating both type I IFN and proinflammatory responses. Despite being predominantly nuclear, IFI16 can unexpectedly sense pathogenic DNA in both the cytoplasm and the nucleus. However, the mechanisms regulating its localization and sensing ability remain uncharacterized. Here, we propose a two-signal model for IFI16 sensing. We first identify an evolutionarily conserved multipartite nuclear localization signal (NLS). Next, using FISH and immunopurification, we demonstrate that IFI16 detects HSV-1 DNA primarily in the nucleus, requiring a functional NLS. Furthermore, we establish a localization-dependent IFN-β induction mediated by IFI16 in response to HSV-1 infection or viral DNA transfection. To identify mechanisms regulating the secondary cytoplasmic localization, we explored IFI16 posttranslation modifications. Combinatorial MS analyses identified numerous acetylations and phosphorylations on endogenous IFI16 in lymphocytes, in which we demonstrate an IFI16-mediated IFN-β response. Importantly, the IFI16 NLS was acetylated in lymphocytes, as well as in macrophages. Mutagenesis and nuclear import assays showed that NLS acetylations promote cytoplasmic localization by inhibiting nuclear import. Additionally, broad-spectrum deacetylase inhibition triggered accumulation of cytoplasmic IFI16, and we identify the acetyltransferase p300 as a regulator of IFI16 localization. Collectively, these studies establish acetylation as a molecular toggle of IFI16 distribution, providing a simple and elegant mechanism by which this versatile sensor detects pathogenic DNA in a localization-dependent manner.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Yan H, et al. RPA nucleic acid-binding properties of IFI16-HIN200. Biochim Biophys Acta. 2008;1784:1087–1097. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
