

Mechanism of membrane perturbation by the HIV-1 gp41 membrane-proximal external region and its modulation by cholesterol
- PMID: 22692008
- PMCID: PMC8796276
- DOI: 10.1016/j.bbamem.2012.06.002
Mechanism of membrane perturbation by the HIV-1 gp41 membrane-proximal external region and its modulation by cholesterol
Abstract
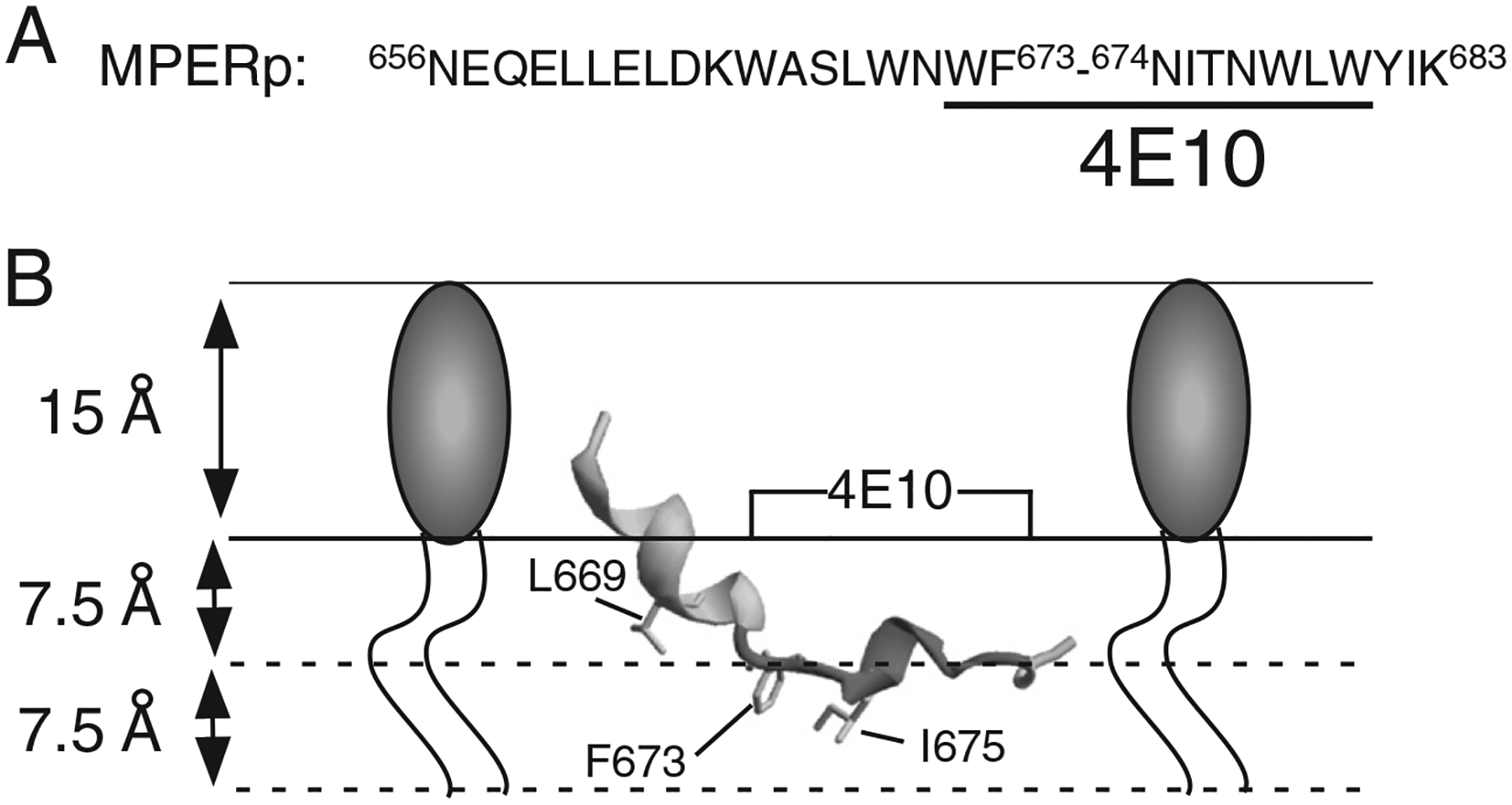
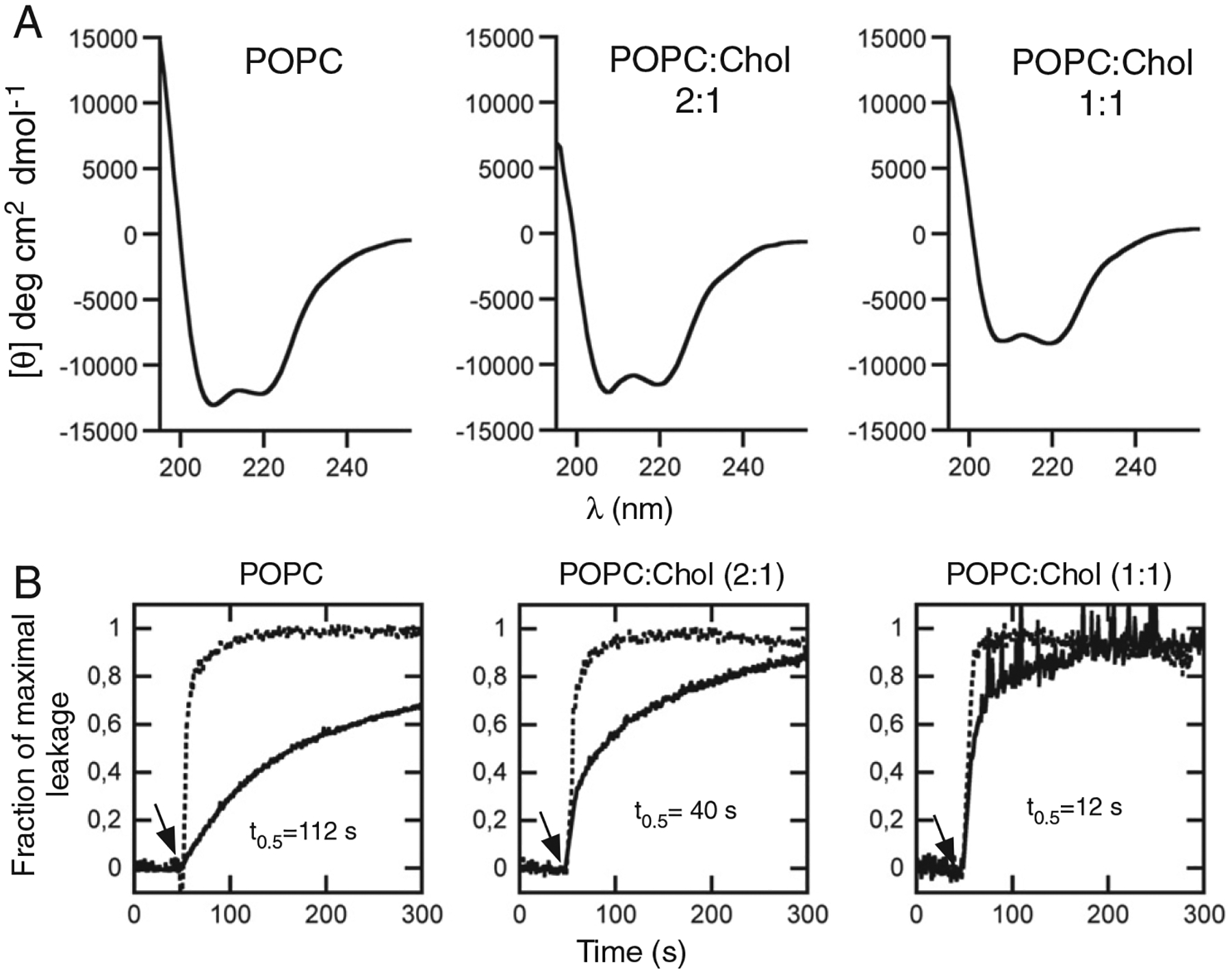
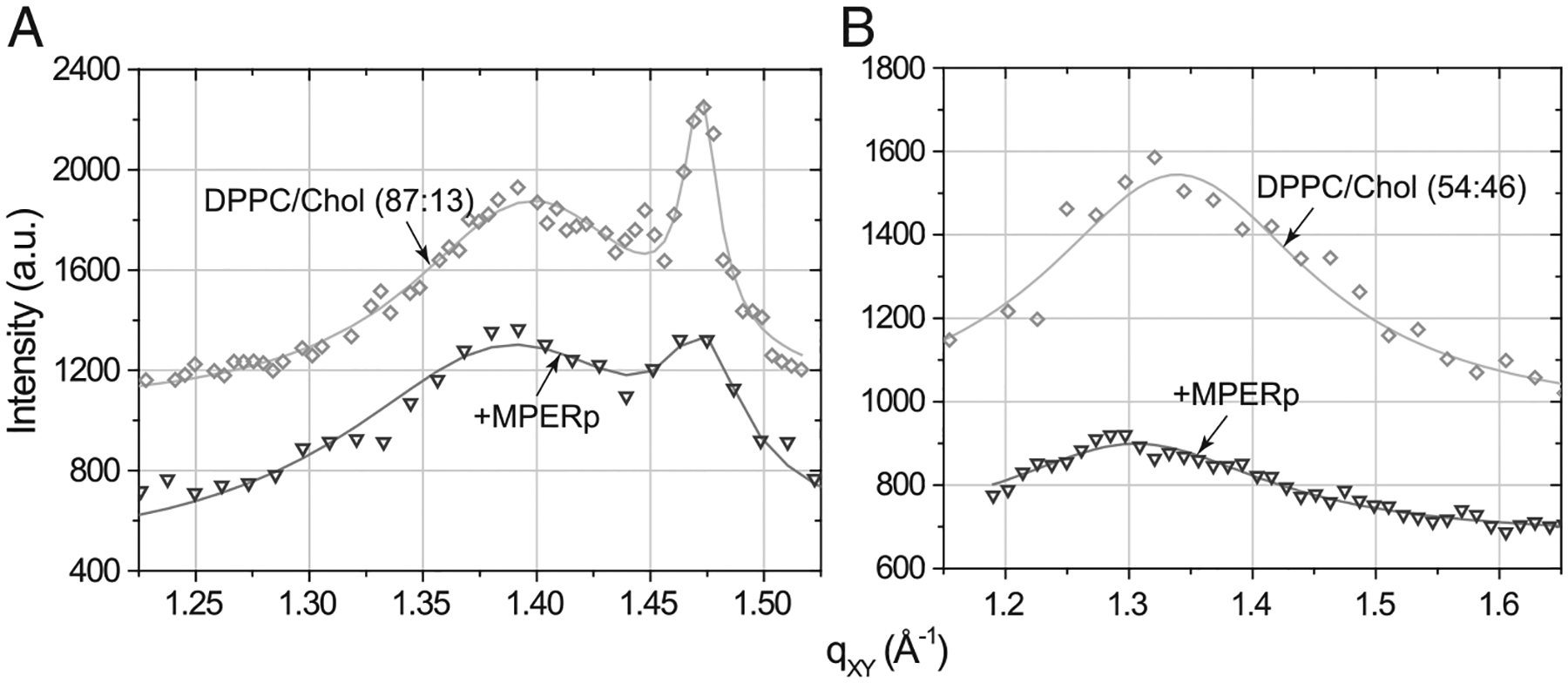
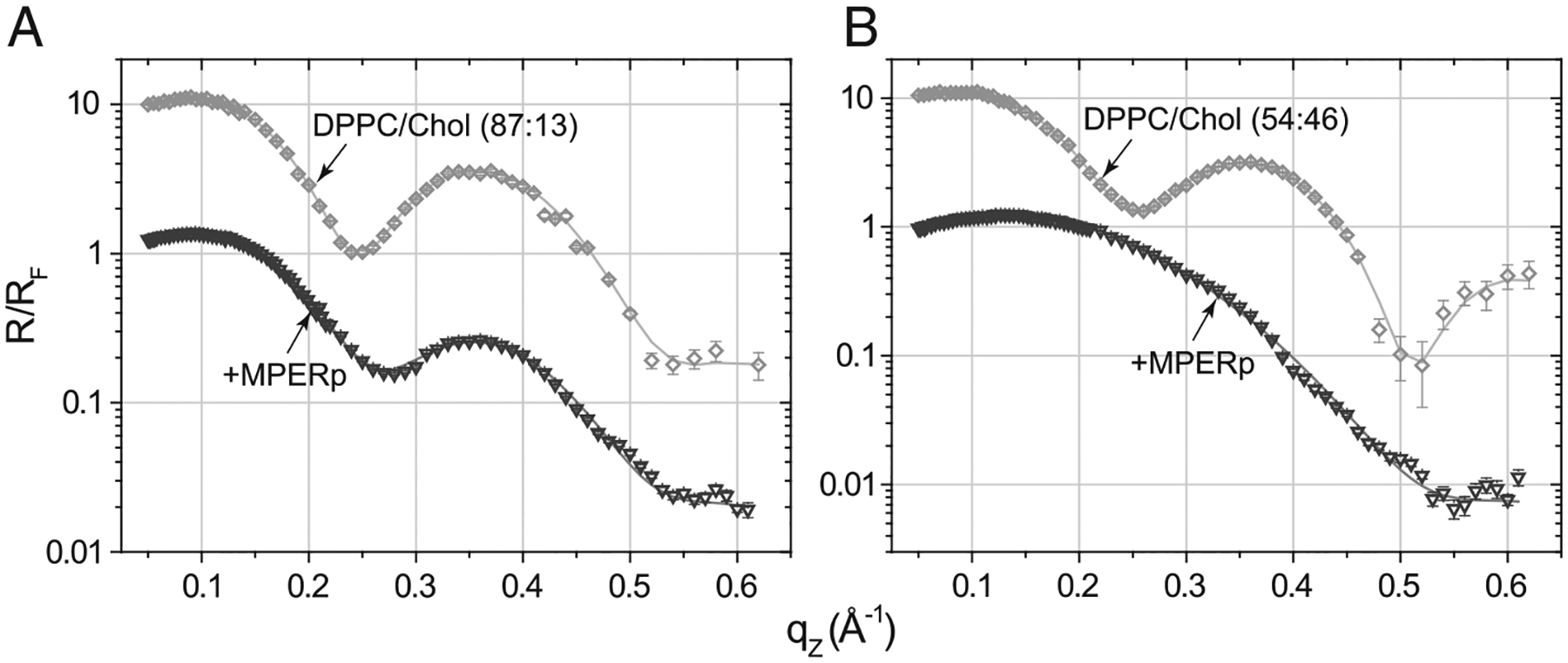
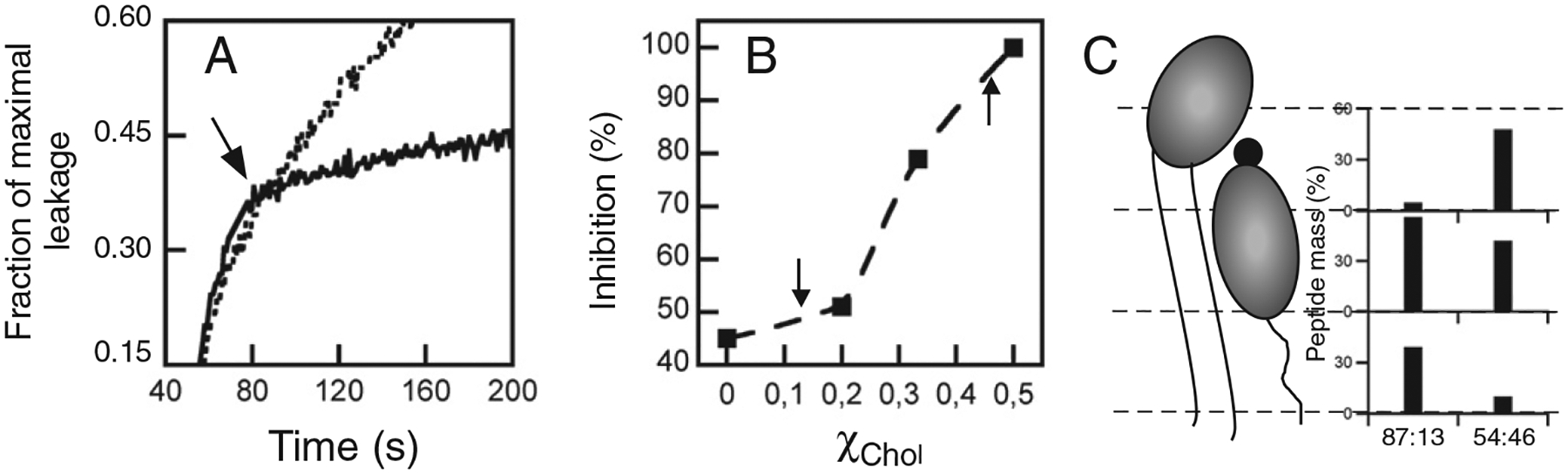
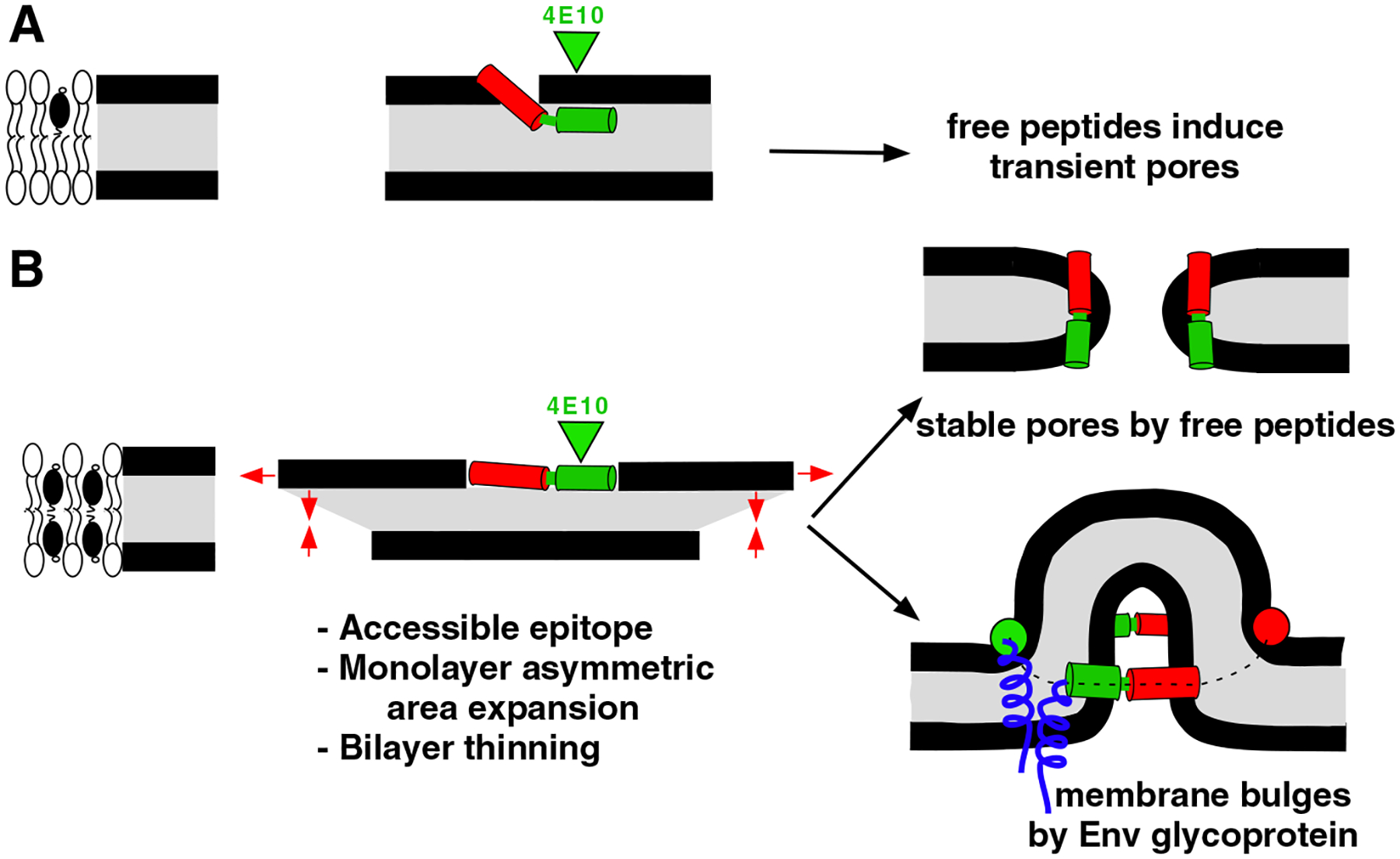
Membrane-activity of the glycoprotein 41 membrane-proximal external region (MPER) is required for HIV-1 membrane fusion. Consequently, its inhibition results in viral neutralization by the antibody 4E10. Previous studies suggested that MPER might act during fusion by locally perturbing the viral membrane, i.e., following a mechanism similar to that proposed for certain antimicrobial peptides. Here, we explore the molecular mechanism of how MPER permeates lipid monolayers containing cholesterol, a main component of the viral envelope, using grazing incidence X-ray diffraction and X-ray reflectivity. Our studies reveal that helical MPER forms lytic pores under conditions not affecting the lateral packing order of lipids. Moreover, we observe an increment of the surface area occupied by MPER helices in cholesterol-enriched membranes, which correlates with an enhancement of the 4E10 epitope accessibility in lipid vesicles. Thus, our data support the view that curvature generation by MPER hydrophobic insertion into the viral membrane is functionally more relevant than lipid packing disruption.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Gallo SA, Finnegan CM, Viard M, Raviv Y, Dimitrov A, Rawat SS, Puri A, Durell S, Blumenthal R, The HIV Env-mediated fusion reaction, Biochim. Biophys. Acta 1614 (2003) 36–50. - PubMed
-
- Peisajovich SG, Shai Y, Viral fusion proteins: multiple regions contribute to membrane fusion, Biochim. Biophys. Acta 1614 (2003) 122–129. - PubMed
-
- Nieva JL, Agirre A, Are fusion peptides a good model to study viral cell fusion? Biochim. Biophys. Acta 1614 (2003) 104–115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
