

Pattern formation during vasculogenesis
- PMID: 22692888
- PMCID: PMC3465733
- DOI: 10.1002/bdrc.21010
Pattern formation during vasculogenesis
Abstract
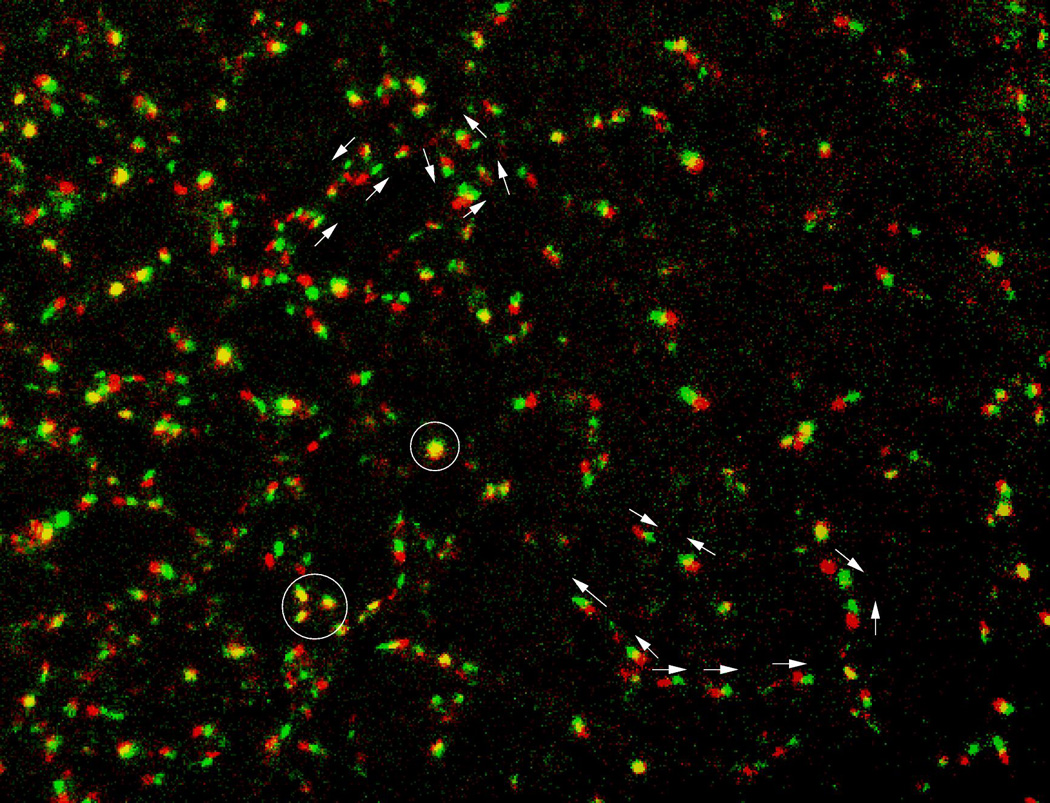
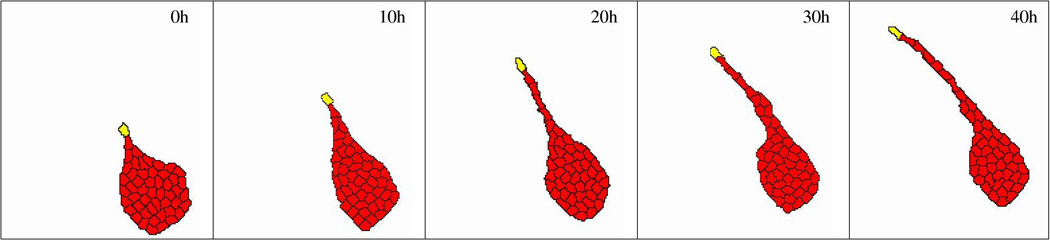
Vasculogenesis, the assembly of the first vascular network, is an intriguing developmental process that yields the first functional organ system of the embryo. In addition to being a fundamental part of embryonic development, vasculogenic processes also have medical importance. To explain the organizational principles behind vascular patterning, we must understand how morphogenesis of tissue level structures can be controlled through cell behavior patterns that, in turn, are determined by biochemical signal transduction processes. Mathematical analyses and computer simulations can help conceptualize how to bridge organizational levels and thus help in evaluating hypotheses regarding the formation of vascular networks. Here, we discuss the ideas that have been proposed to explain the formation of the first vascular pattern: cell motility guided by extracellular matrix alignment (contact guidance), chemotaxis guided by paracrine and autocrine morphogens, and sprouting guided by cell-cell contacts.
Copyright © 2012 Wiley Periodicals, Inc.
Figures





References
-
- Alber M, Chen N, Glimm T, Lushnikov PM. Multiscale dynamics of biological cells with chemotactic interactions: from a discrete stochastic model to a continuous description. Phys Rev E Stat Nonlin Soft Matter Phys. 2006;73:051901. - PubMed
-
- Alt W. Biased random walk models for chemotaxis and related diffusion approximations. J Math Biol. 1980;9:147–177. - PubMed
-
- Ambler CA, Nowicki JL, Burke AC, Bautch VL. Assembly of trunk and limb blood vessels involves extensive migration and vasculogenesis of somite-derived angioblasts. Dev Biol. 2001;234:352–364. - PubMed
-
- Anderson A, Chaplain M. Continuous and discrete mathematical-models of tumor-induced angiogenesis. Bulletin of mathematical biology. 1998;60:857–899. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
