

From sensor data to animal behaviour: an oystercatcher example
- PMID: 22693586
- PMCID: PMC3365100
- DOI: 10.1371/journal.pone.0037997
From sensor data to animal behaviour: an oystercatcher example
Abstract
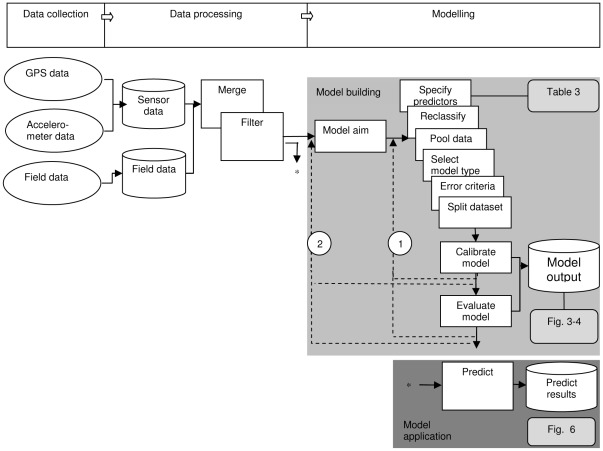
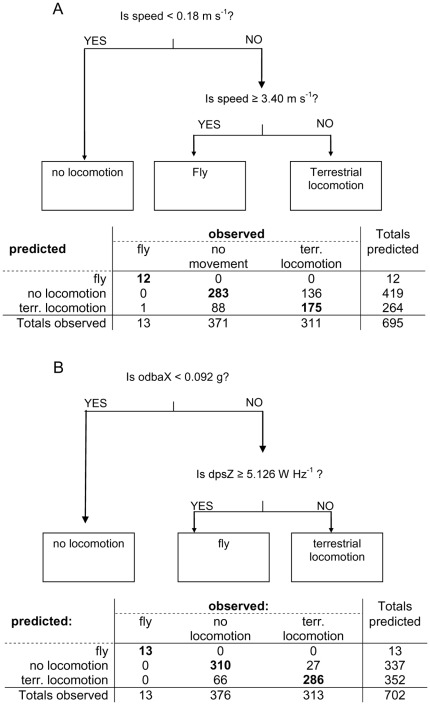
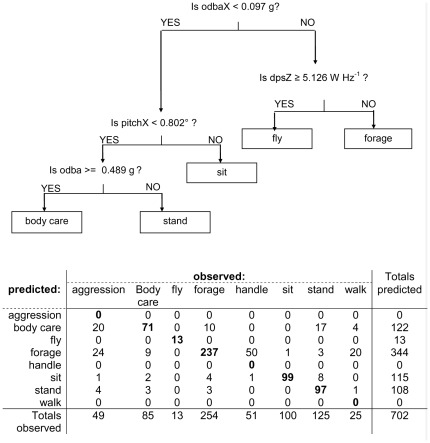
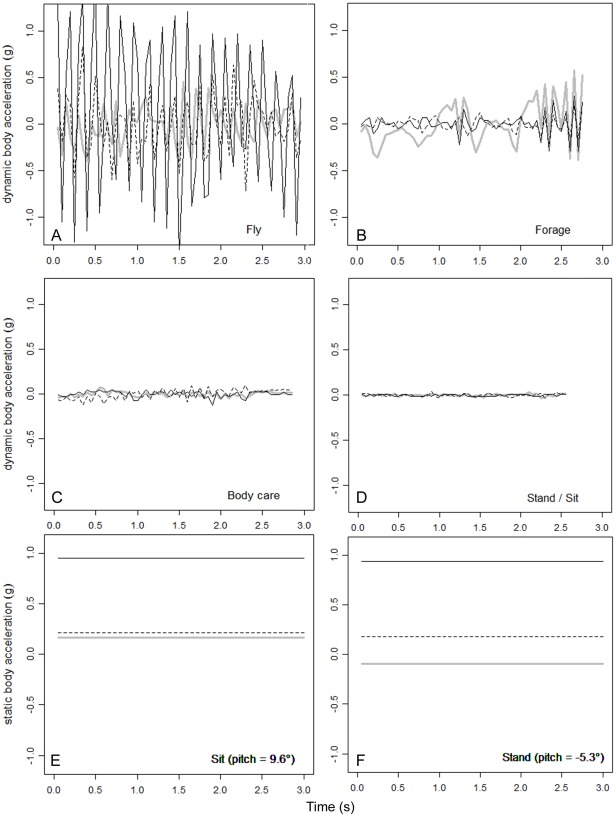
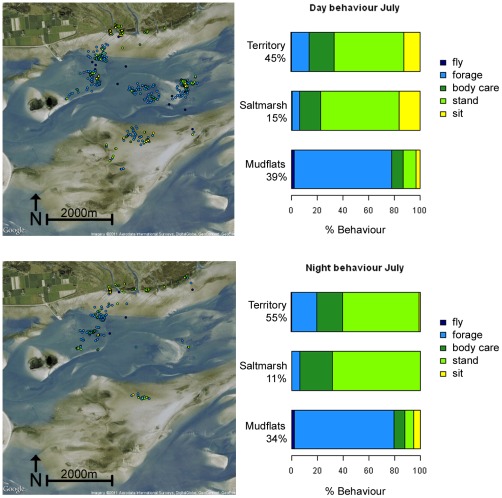
Animal-borne sensors enable researchers to remotely track animals, their physiological state and body movements. Accelerometers, for example, have been used in several studies to measure body movement, posture, and energy expenditure, although predominantly in marine animals. In many studies, behaviour is often inferred from expert interpretation of sensor data and not validated with direct observations of the animal. The aim of this study was to derive models that could be used to classify oystercatcher (Haematopus ostralegus) behaviour based on sensor data. We measured the location, speed, and tri-axial acceleration of three oystercatchers using a flexible GPS tracking system and conducted simultaneous visual observations of the behaviour of these birds in their natural environment. We then used these data to develop three supervised classification trees of behaviour and finally applied one of the models to calculate time-activity budgets. The model based on accelerometer data developed to classify three behaviours (fly, terrestrial locomotion, and no movement) was much more accurate (cross-validation error = 0.14) than the model based on GPS-speed alone (cross-validation error = 0.35). The most parsimonious acceleration model designed to classify eight behaviours could distinguish five: fly, forage, body care, stand, and sit (cross-validation error = 0.28); other behaviours that were observed, such as aggression or handling of prey, could not be distinguished. Model limitations and potential improvements are discussed. The workflow design presented in this study can facilitate model development, be adapted to a wide range of species, and together with the appropriate measurements, can foster the study of behaviour and habitat use of free living animals throughout their annual routine.
Conflict of interest statement
Figures

References
-
- Norberg RA. An ecological theory on foraging time and energetics and choice of optimal food-searching method. J Anim Ecol. 1977;46:511–529.
-
- Ropert-Coudert Y, Grémillet D, Kato A, Ryan PG, Naito Y, et al. A fine-scale time budget of Cape gannets provides insights into the foraging strategies of coastal seabirds. Anim Behav. 2004;67:985–992.
-
- Aviles JM. Common cranes Grus grus and habitat management in holm oak dehesas of Spain. Biodivers Conserv. 2004;13:2015–2025.
-
- Bograd SJ, Block BA, Costa DP, Godley BJ. Biologging technologies: new tools for conservation. Introduction. Endangered Species Research. 2010;10:1–7.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
