

Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts
- PMID: 22699508
- PMCID: PMC3390844
- DOI: 10.1073/pnas.1203287109
Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts
Abstract
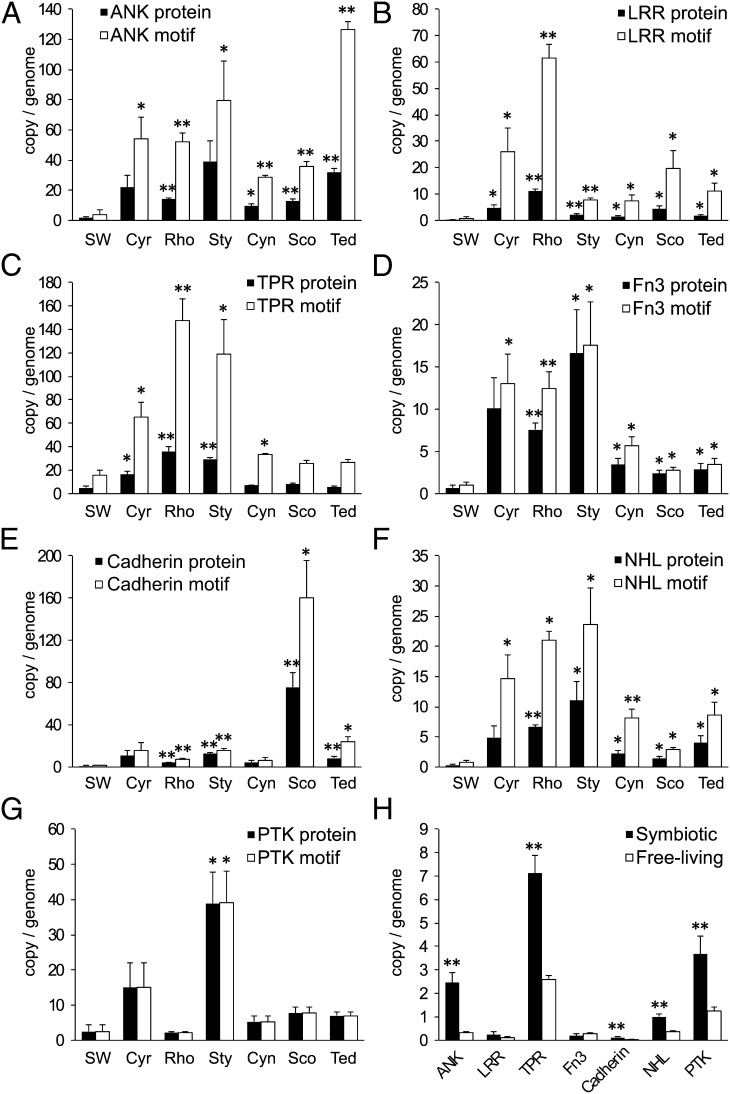
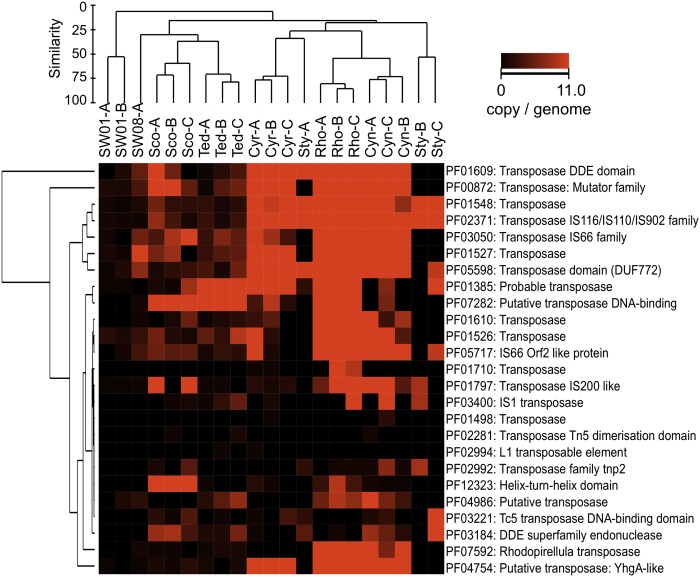
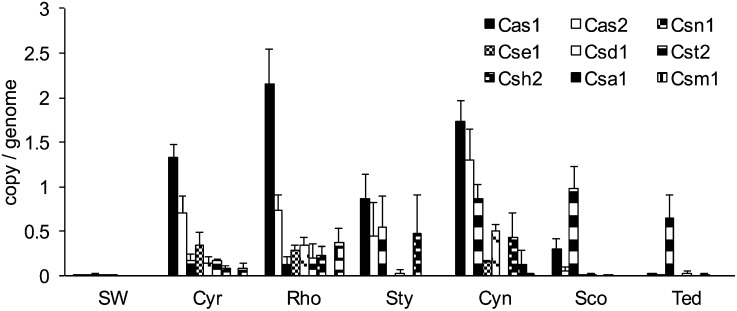
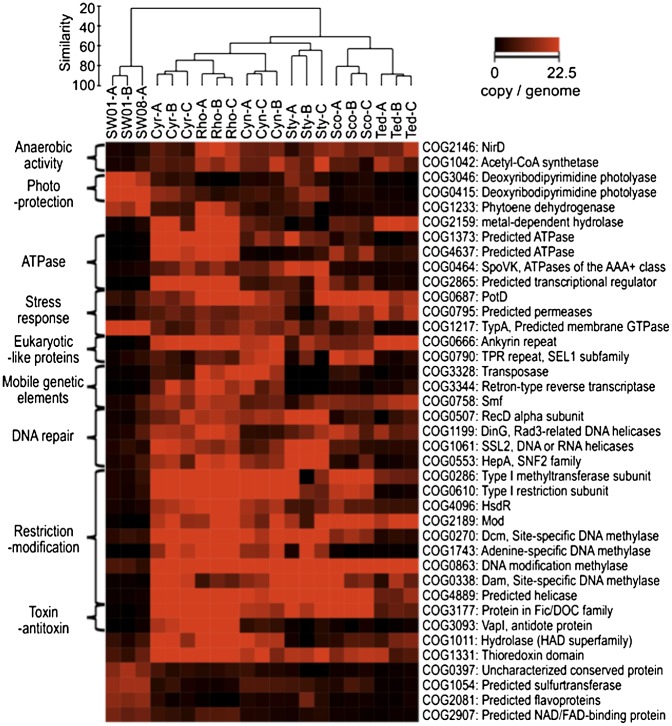
Microorganisms often form symbiotic relationships with eukaryotes, and the complexity of these relationships can range from those with one single dominant symbiont to associations with hundreds of symbiont species. Microbial symbionts occupying equivalent niches in different eukaryotic hosts may share functional aspects, and convergent genome evolution has been reported for simple symbiont systems in insects. However, for complex symbiont communities, it is largely unknown how prevalent functional equivalence is and whether equivalent functions are conducted by evolutionarily convergent mechanisms. Sponges represent an evolutionarily divergent group of species with common physiological and ecological traits. They also host complex communities of microbial symbionts and thus are the ideal model to test whether functional equivalence and evolutionary convergence exist in complex symbiont communities across phylogenetically divergent hosts. Here we use a sampling design to determine the phylogenetic and functional profiles of microbial communities associated with six sponge species. We identify common functions in the six microbiomes, demonstrating the existence of functional equivalence. These core functions are consistent with our current understanding of the biological and ecological roles of sponge-associated microorganisms and also provide insight into symbiont functions. Importantly, core functions also are provided in each sponge species by analogous enzymes and biosynthetic pathways. Moreover, the abundance of elements involved in horizontal gene transfer suggests their key roles in the genomic evolution of symbionts. Our data thus demonstrate evolutionary convergence in complex symbiont communities and reveal the details and mechanisms that underpin the process.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Diversity, structure and convergent evolution of the global sponge microbiome.Nat Commun. 2016 Jun 16;7:11870. doi: 10.1038/ncomms11870. Nat Commun. 2016. PMID: 27306690 Free PMC article.
-
Metagenomic Analysis of Genes Encoding Nutrient Cycling Pathways in the Microbiota of Deep-Sea and Shallow-Water Sponges.Mar Biotechnol (NY). 2016 Dec;18(6):659-671. doi: 10.1007/s10126-016-9725-5. Epub 2016 Nov 7. Mar Biotechnol (NY). 2016. PMID: 27819120
-
Dynamic microbiome diversity shaping the adaptation of sponge holobionts in coastal waters.Microbiol Spectr. 2024 Nov 5;12(11):e0144824. doi: 10.1128/spectrum.01448-24. Epub 2024 Oct 14. Microbiol Spectr. 2024. PMID: 39400157 Free PMC article.
-
An environmental genomics perspective on the diversity and function of marine sponge-associated microbiota.Curr Opin Microbiol. 2007 Jun;10(3):215-20. doi: 10.1016/j.mib.2007.05.012. Epub 2007 Jun 15. Curr Opin Microbiol. 2007. PMID: 17574904 Review.
-
Sponge-microbe symbioses: recent advances and new directions.Adv Mar Biol. 2012;62:57-111. doi: 10.1016/B978-0-12-394283-8.00002-3. Adv Mar Biol. 2012. PMID: 22664121 Review.
Cited by
-
Coral reef invertebrate microbiomes correlate with the presence of photosymbionts.ISME J. 2013 Jul;7(7):1452-8. doi: 10.1038/ismej.2012.172. Epub 2013 Jan 10. ISME J. 2013. PMID: 23303372 Free PMC article.
-
Sample Processing Impacts the Viability and Cultivability of the Sponge Microbiome.Front Microbiol. 2016 Apr 12;7:499. doi: 10.3389/fmicb.2016.00499. eCollection 2016. Front Microbiol. 2016. PMID: 27242673 Free PMC article.
-
The different potential of sponge bacterial symbionts in N₂ release indicated by the phylogenetic diversity and abundance analyses of denitrification genes, nirK and nosZ.PLoS One. 2013 Jun 10;8(6):e65142. doi: 10.1371/journal.pone.0065142. Print 2013. PLoS One. 2013. PMID: 23762300 Free PMC article.
-
Phylogenetic beta diversity in bacterial assemblages across ecosystems: deterministic versus stochastic processes.ISME J. 2013 Jul;7(7):1310-21. doi: 10.1038/ismej.2013.30. Epub 2013 Feb 28. ISME J. 2013. PMID: 23446837 Free PMC article.
-
Diversity, structure and convergent evolution of the global sponge microbiome.Nat Commun. 2016 Jun 16;7:11870. doi: 10.1038/ncomms11870. Nat Commun. 2016. PMID: 27306690 Free PMC article.
References
-
- Serbus LR, Casper-Lindley C, Landmann F, Sullivan W. The genetics and cell biology of Wolbachia-host interactions. Annu Rev Genet. 2008;42:683–707. - PubMed
-
- McFall-Ngai M. Host-microbe symbiosis: The squid-Vibrio association—a naturally occurring, experimental model of animal/bacterial partnerships. Adv Exp Med Biol. 2008;635:102–112. - PubMed
-
- Marchesi JR. Prokaryotic and eukaryotic diversity of the human gut. Adv Appl Microbiol. 2010;72:43–62. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
