

Function, targets, and evolution of Caenorhabditis elegans piRNAs
- PMID: 22700655
- PMCID: PMC3951736
- DOI: 10.1126/science.1220952
Function, targets, and evolution of Caenorhabditis elegans piRNAs
Abstract
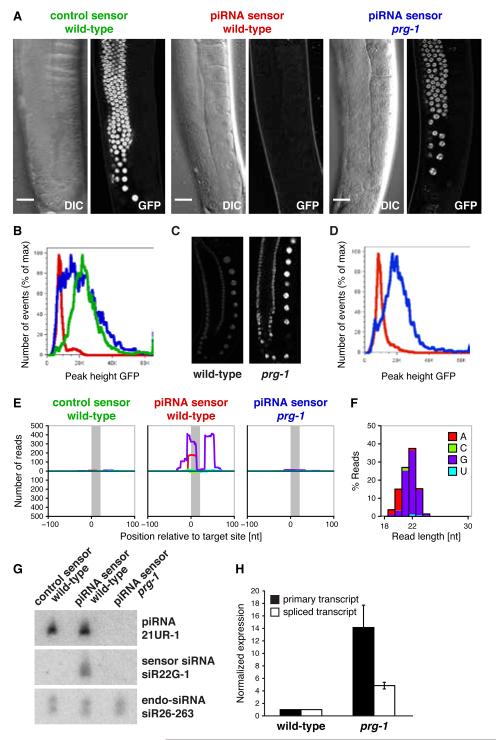
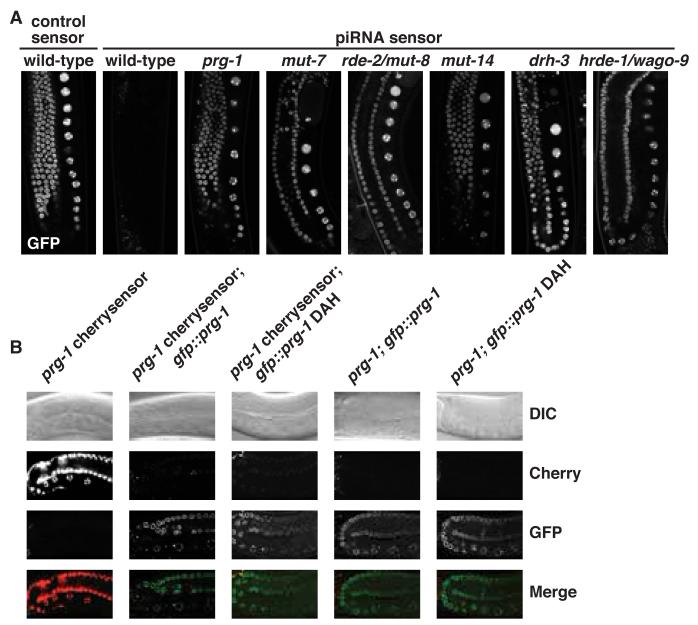
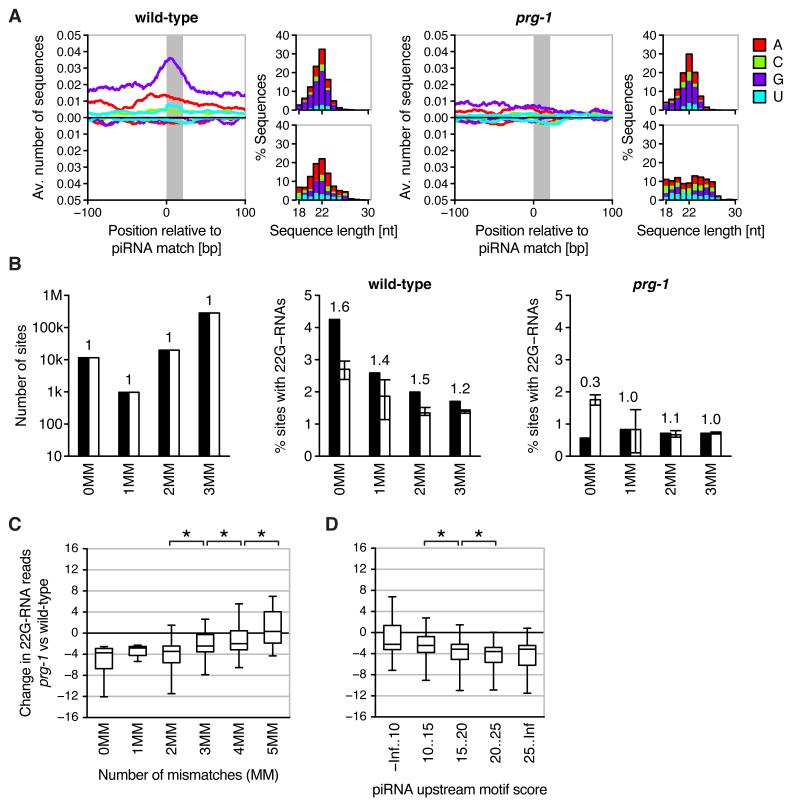
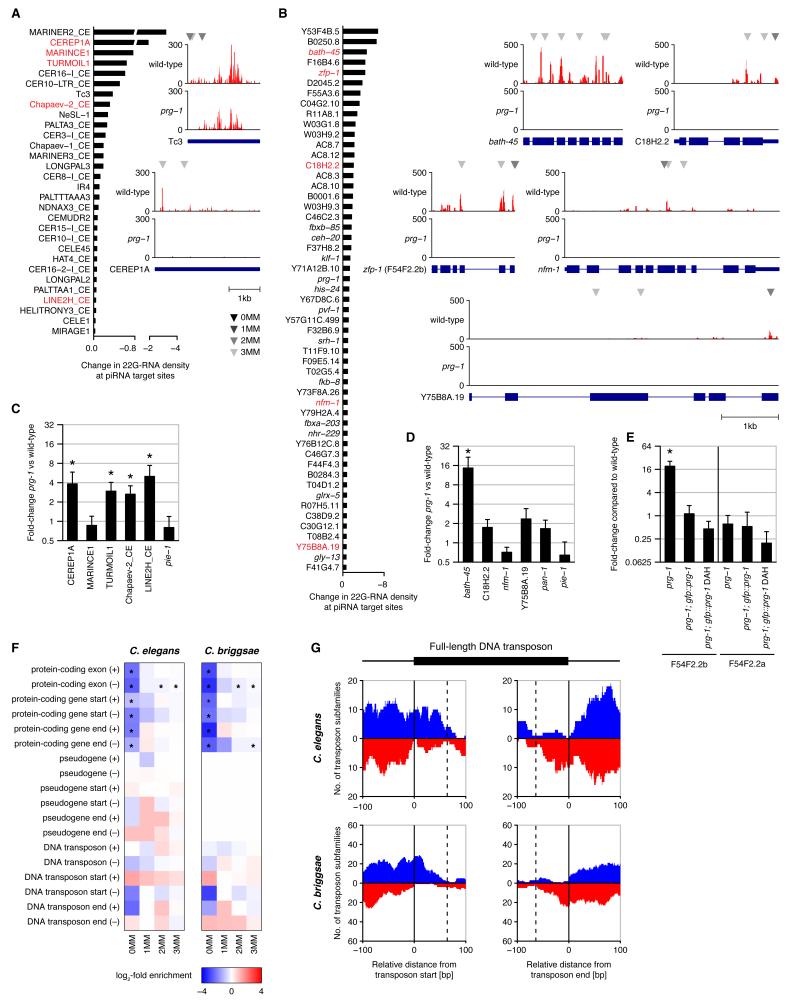
Piwi-interacting RNAs (piRNAs) are small RNAs required to maintain germline integrity and fertility, but their mechanism of action is poorly understood. Here we demonstrate that Caenorhabditis elegans piRNAs silence transcripts in trans through imperfectly complementary sites. Target silencing is independent of Piwi endonuclease activity or "slicing." Instead, piRNAs initiate a localized secondary endogenous small interfering RNA (endo-siRNA) response. Endogenous protein-coding gene and transposon transcripts exhibit Piwi-dependent endo-siRNAs at sites complementary to piRNAs and are derepressed in Piwi mutants. Genomic loci of piRNA biogenesis are depleted of protein-coding genes and tend to overlap the start and end of transposons in sense and antisense, respectively. Our data suggest that nematode piRNA clusters are evolving to generate piRNAs against active mobile elements. Thus, piRNAs provide heritable, sequence-specific triggers for RNA interference in C. elegans.
Figures
Comment in
-
Small RNAs: piRNA surveillance in the C. elegans germline.Nat Rev Genet. 2012 Jul 10;13(8):518-9. doi: 10.1038/nrg3289. Nat Rev Genet. 2012. PMID: 22777129 No abstract available.
-
Small RNAs: transmitting silence through generations.Nat Rev Mol Cell Biol. 2012 Jul 23;13(8):477. doi: 10.1038/nrm3406. Nat Rev Mol Cell Biol. 2012. PMID: 22820881 No abstract available.
-
Molecular biology. Outsourcing genome protection.Science. 2012 Aug 3;337(6094):529-30. doi: 10.1126/science.1227095. Science. 2012. PMID: 22859473 No abstract available.
References
-
- Ruby J, et al. Large-Scale Sequencing Reveals 21U-RNAs and Additional MicroRNAs and Endogenous siRNAs in C. elegans. Cell. 2006;127:1193–1207. - PubMed
-
- Brennecke J, et al. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell. 2007;128:1089–1103. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
