

Mutations in the GlyT2 gene (SLC6A5) are a second major cause of startle disease
- PMID: 22700964
- PMCID: PMC3436555
- DOI: 10.1074/jbc.M112.372094
Mutations in the GlyT2 gene (SLC6A5) are a second major cause of startle disease
Abstract
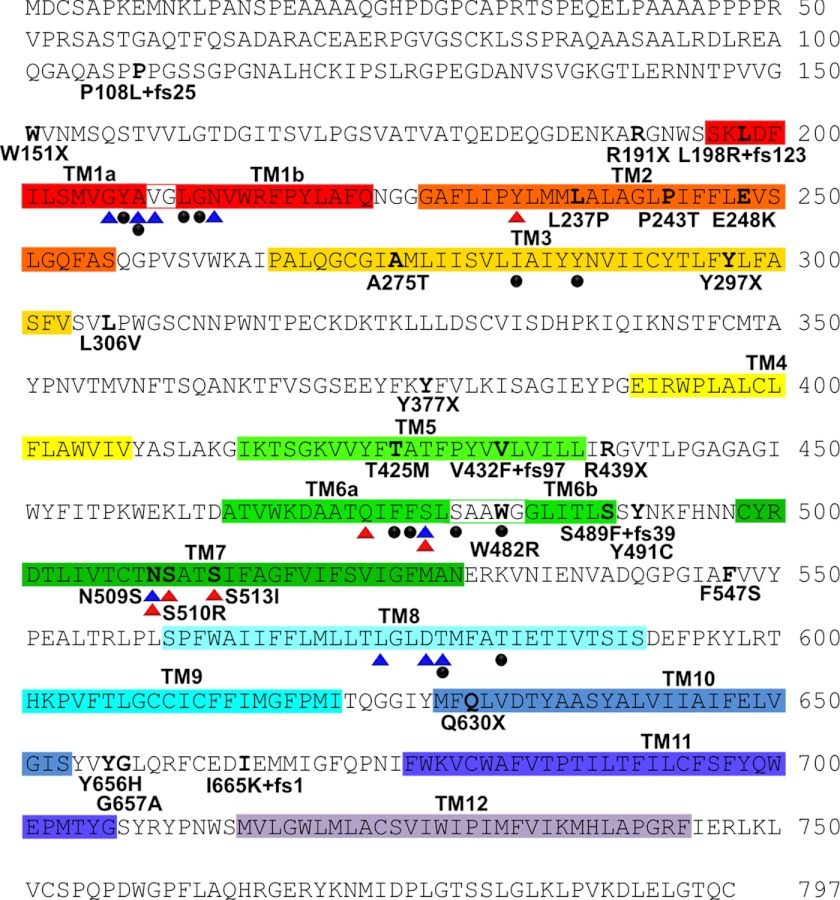
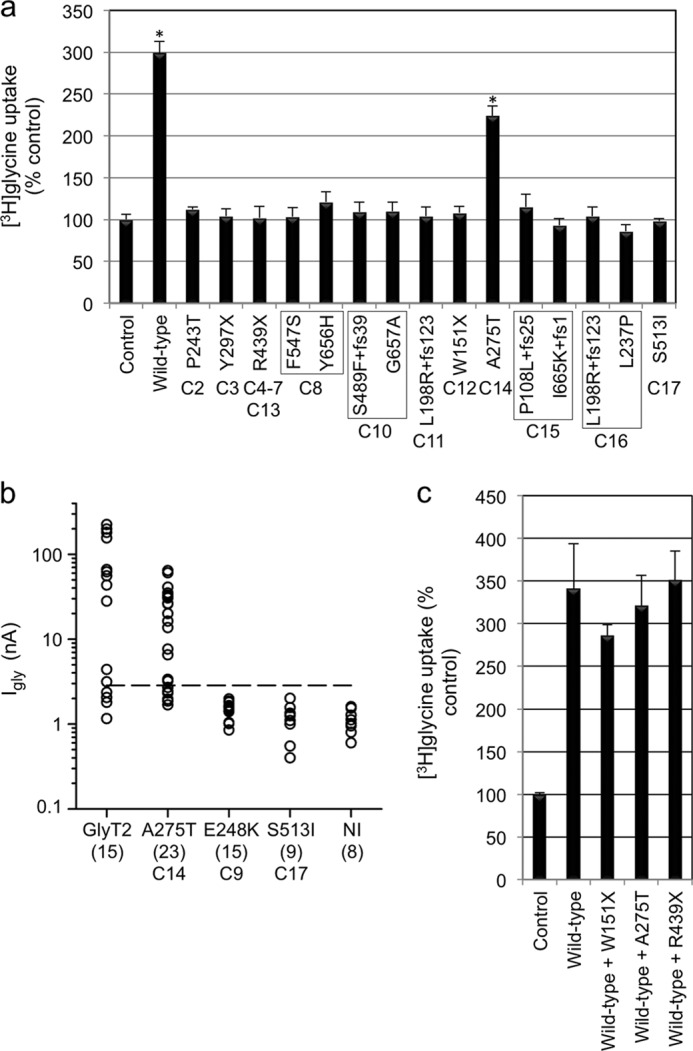
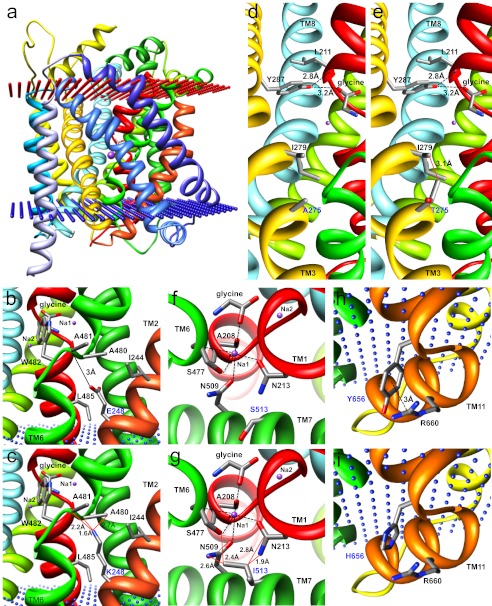
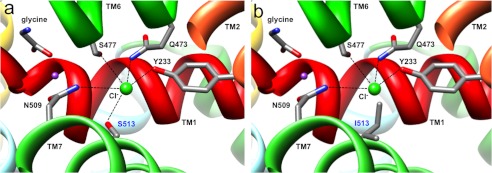
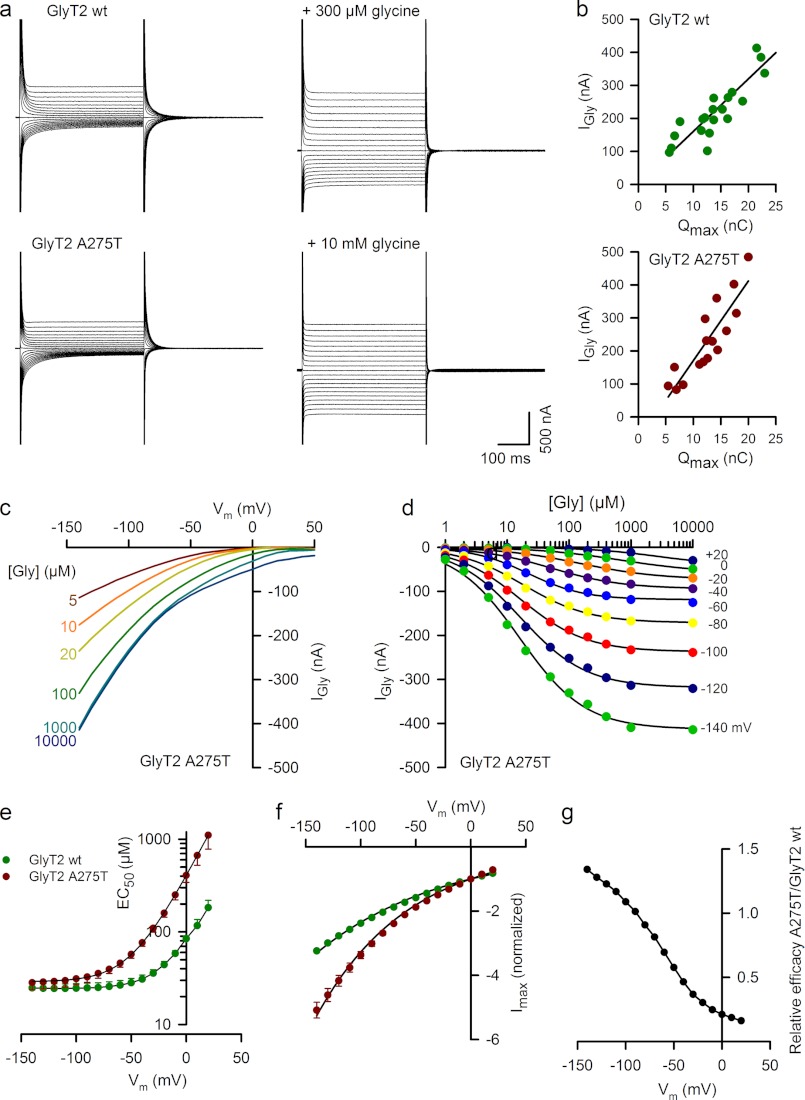
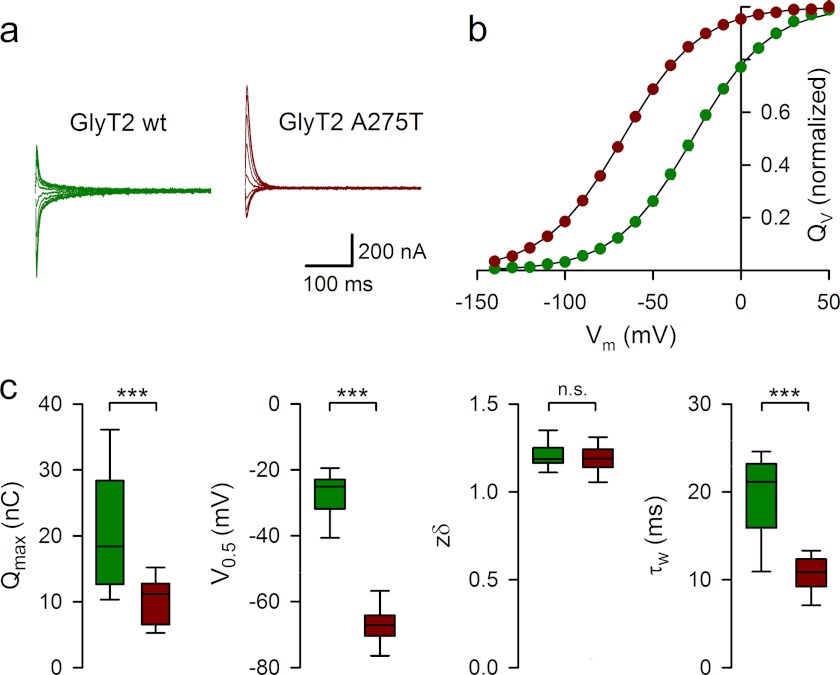
Hereditary hyperekplexia or startle disease is characterized by an exaggerated startle response, evoked by tactile or auditory stimuli, leading to hypertonia and apnea episodes. Missense, nonsense, frameshift, splice site mutations, and large deletions in the human glycine receptor α1 subunit gene (GLRA1) are the major known cause of this disorder. However, mutations are also found in the genes encoding the glycine receptor β subunit (GLRB) and the presynaptic Na(+)/Cl(-)-dependent glycine transporter GlyT2 (SLC6A5). In this study, systematic DNA sequencing of SLC6A5 in 93 new unrelated human hyperekplexia patients revealed 20 sequence variants in 17 index cases presenting with homozygous or compound heterozygous recessive inheritance. Five apparently unrelated cases had the truncating mutation R439X. Genotype-phenotype analysis revealed a high rate of neonatal apneas and learning difficulties associated with SLC6A5 mutations. From the 20 SLC6A5 sequence variants, we investigated glycine uptake for 16 novel mutations, confirming that all were defective in glycine transport. Although the most common mechanism of disrupting GlyT2 function is protein truncation, new pathogenic mechanisms included splice site mutations and missense mutations affecting residues implicated in Cl(-) binding, conformational changes mediated by extracellular loop 4, and cation-π interactions. Detailed electrophysiology of mutation A275T revealed that this substitution results in a voltage-sensitive decrease in glycine transport caused by lower Na(+) affinity. This study firmly establishes the combination of missense, nonsense, frameshift, and splice site mutations in the GlyT2 gene as the second major cause of startle disease.
Figures
References
-
- Nigro M. A., Lim H. C. (1992) Hyperekplexia and sudden neonatal death. Pediatr. Neurol. 8, 221–225 - PubMed
-
- Vigevano F., Di Capua M., Dalla Bernardina B. (1989) Startle disease. An avoidable cause of sudden infant death. Lancet 1, 216. - PubMed
-
- Bakker M. J., van Dijk J. G., van den Maagdenberg A. M., Tijssen M. A. (2006) Startle syndromes. Lancet Neurol. 5, 513–524 - PubMed
-
- Thomas R. H., Stephenson J. B., Harvey R. J., Rees M. I. (2010) Hyperekplexia. Stiffness, startle and syncope. J. Ped. Neurol. 8, 11–14
-
- Bakker M. J., Peeters E. A., Tijssen M. A. (2009) Clonazepam is an effective treatment for hyperekplexia due to a SLC6A5 (GlyT2) mutation. Mov. Disord. 24, 1852–1854 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
