

Structural basis for the interaction of a hexameric replicative helicase with the regulatory subunit of human DNA polymerase α-primase
- PMID: 22700977
- PMCID: PMC3411022
- DOI: 10.1074/jbc.M112.363655
Structural basis for the interaction of a hexameric replicative helicase with the regulatory subunit of human DNA polymerase α-primase
Abstract
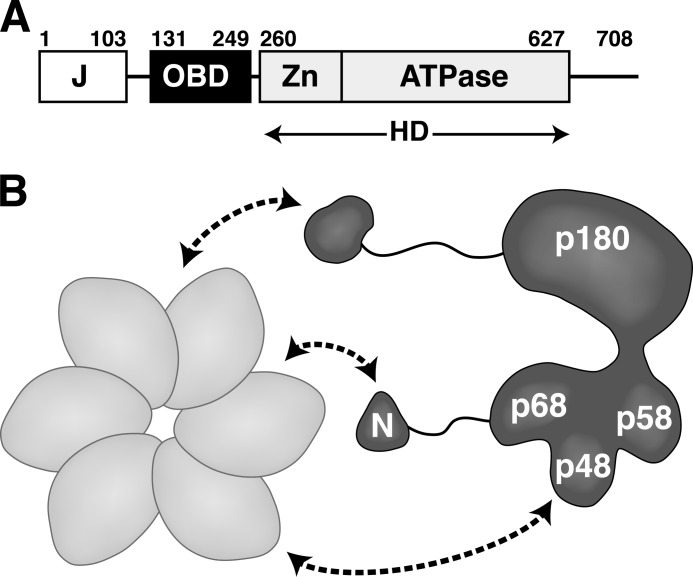
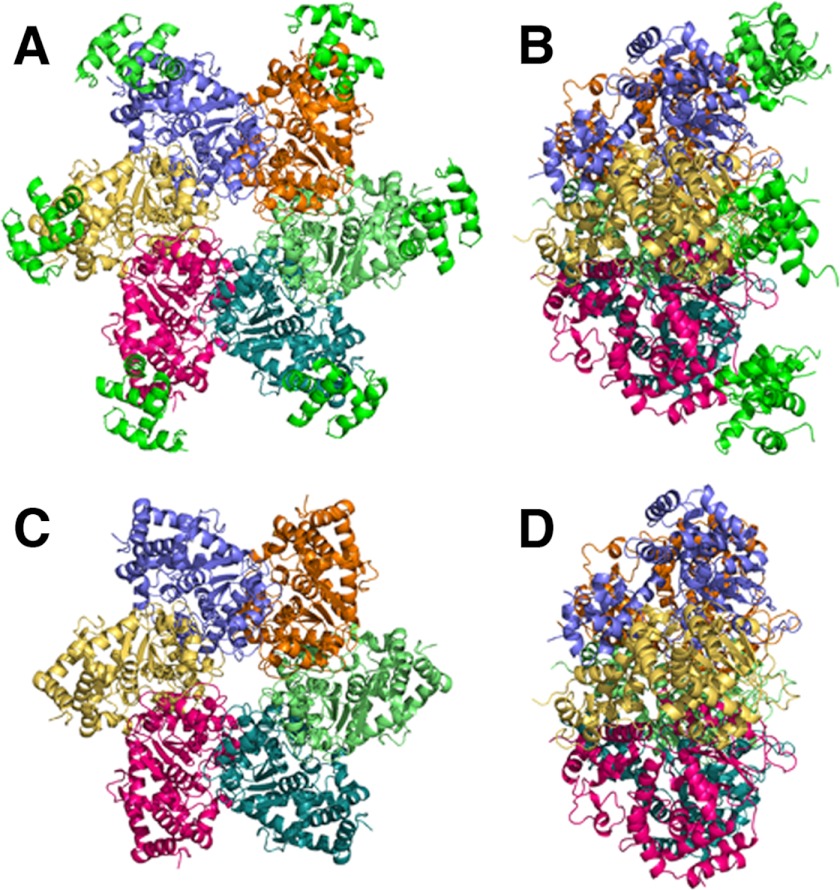
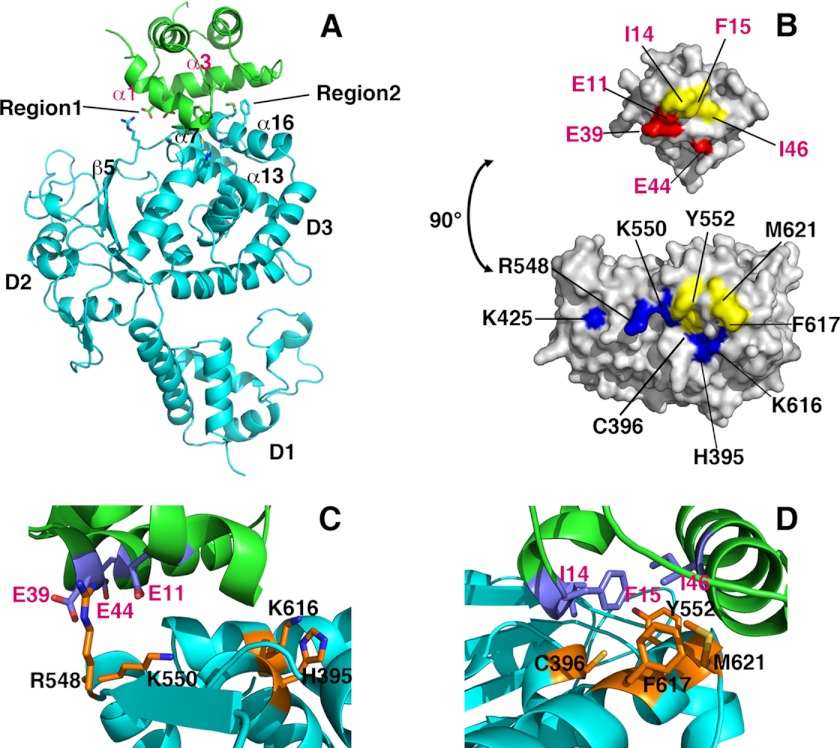
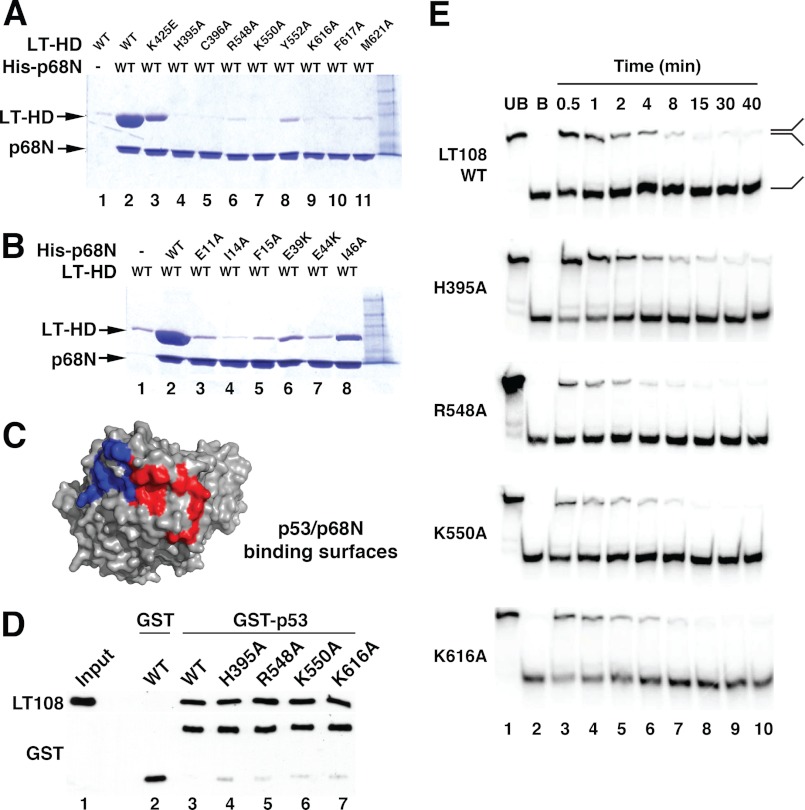
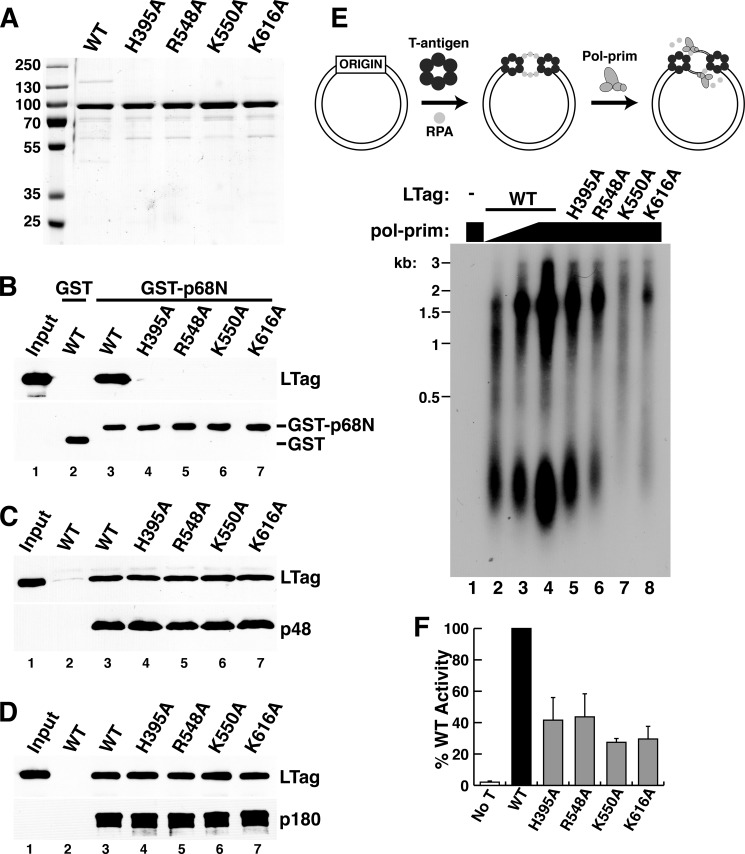
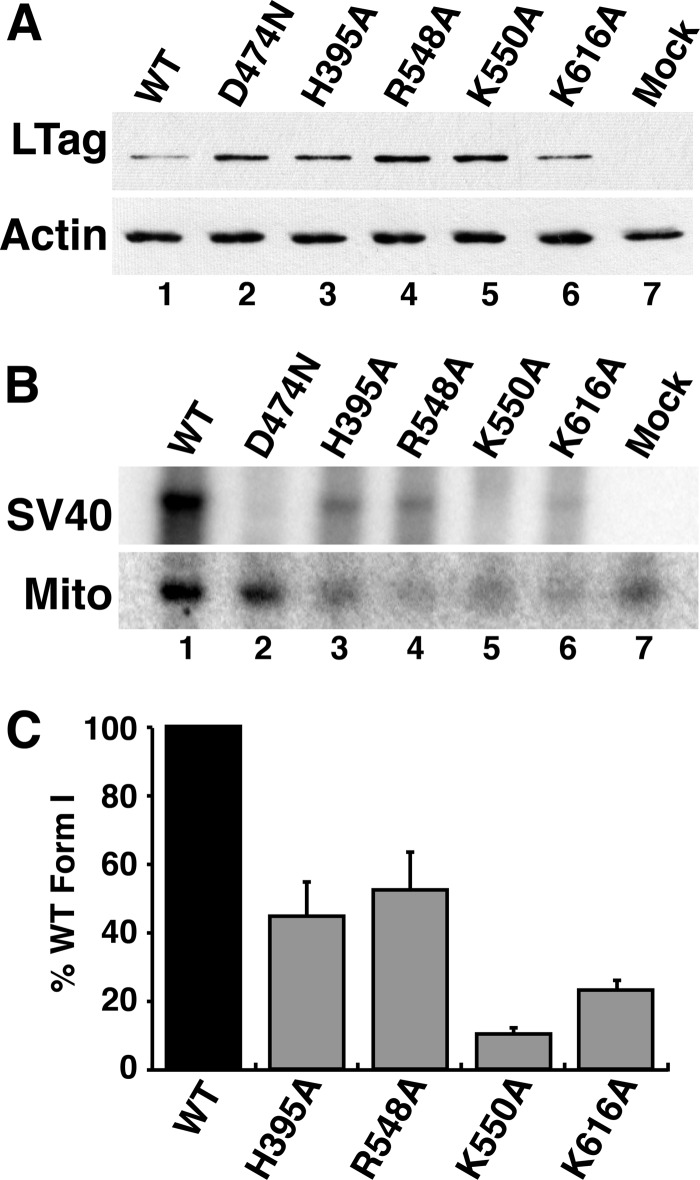
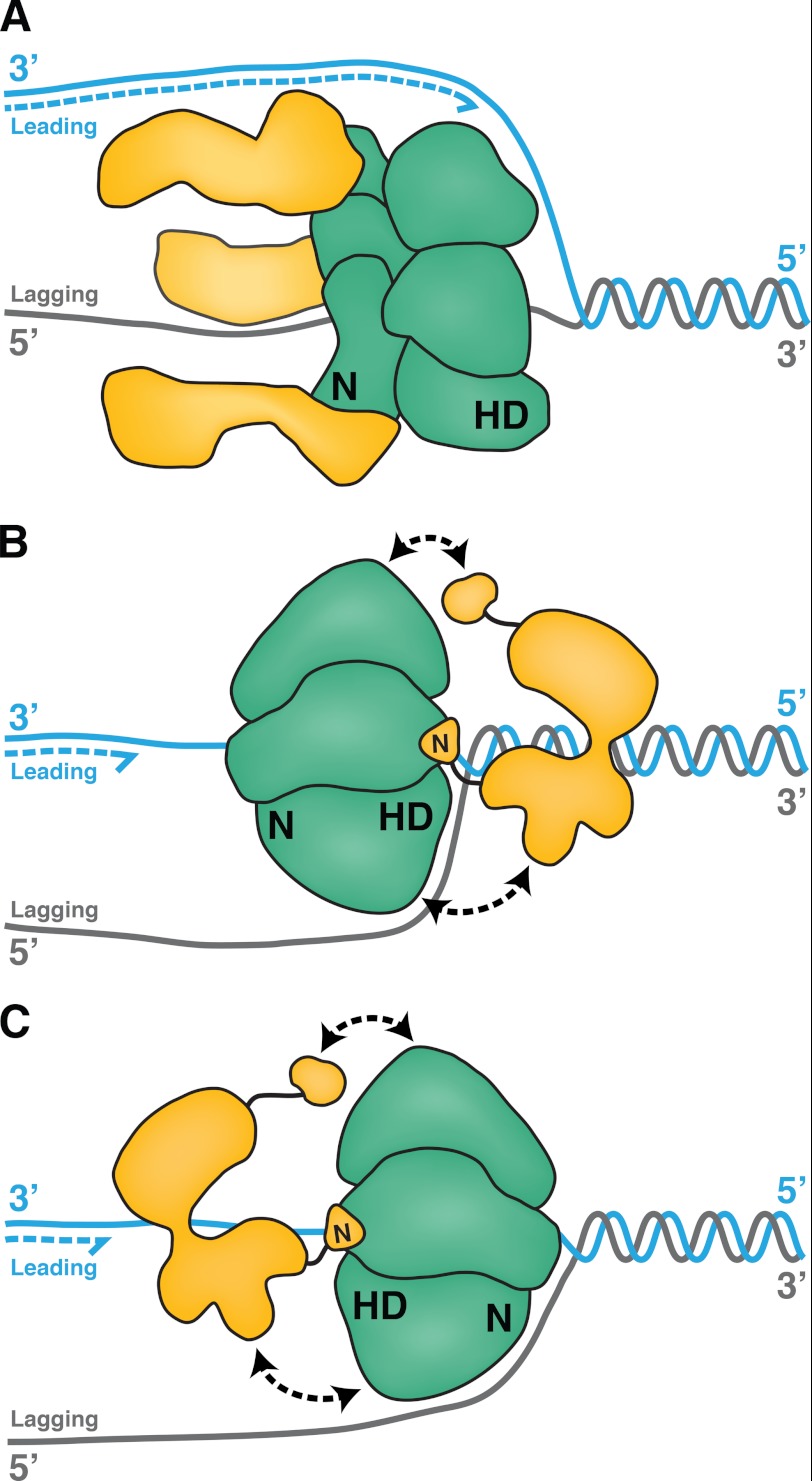
DNA polymerase α-primase (Pol-prim) plays an essential role in eukaryotic DNA replication, initiating synthesis of the leading strand and of each Okazaki fragment on the lagging strand. Pol-prim is composed of a primase heterodimer that synthesizes an RNA primer, a DNA polymerase subunit that extends the primer, and a regulatory B-subunit (p68) without apparent enzymatic activity. Pol-prim is thought to interact with eukaryotic replicative helicases, forming a dynamic multiprotein assembly that displays primosome activity. At least three subunits of Pol-prim interact physically with the hexameric replicative helicase SV40 large T antigen, constituting a simple primosome that is active in vitro. However, structural understanding of these interactions and their role in viral chromatin replication in vivo remains incomplete. Here, we report the detailed large T antigen-p68 interface, as revealed in a co-crystal structure and validated by site-directed mutagenesis, and we demonstrate its functional importance in activating the SV40 primosome in cell-free reactions with purified Pol-prim, as well as in monkey cells in vivo.
Figures
Similar articles
-
Structure of a DNA polymerase alpha-primase domain that docks on the SV40 helicase and activates the viral primosome.J Biol Chem. 2010 May 28;285(22):17112-22. doi: 10.1074/jbc.M110.116830. Epub 2010 Mar 16. J Biol Chem. 2010. PMID: 20234039 Free PMC article.
-
A specific docking site for DNA polymerase {alpha}-primase on the SV40 helicase is required for viral primosome activity, but helicase activity is dispensable.J Biol Chem. 2010 Oct 22;285(43):33475-33484. doi: 10.1074/jbc.M110.156240. Epub 2010 Aug 3. J Biol Chem. 2010. PMID: 20685648 Free PMC article.
-
Protein-protein interactions of the primase subunits p58 and p48 with simian virus 40 T antigen are required for efficient primer synthesis in a cell-free system.J Biol Chem. 2000 Jun 9;275(23):17328-37. doi: 10.1074/jbc.M000717200. J Biol Chem. 2000. PMID: 10747950
-
The DNA polymerase alpha-primase complex: multiple functions and interactions.ScientificWorldJournal. 2003 Mar 17;3:21-33. doi: 10.1100/tsw.2003.05. ScientificWorldJournal. 2003. PMID: 12806117 Free PMC article. Review.
-
The Pol α-primase complex.Subcell Biochem. 2012;62:157-69. doi: 10.1007/978-94-007-4572-8_9. Subcell Biochem. 2012. PMID: 22918585 Review.
Cited by
-
ATM and ATR activities maintain replication fork integrity during SV40 chromatin replication.PLoS Pathog. 2013;9(4):e1003283. doi: 10.1371/journal.ppat.1003283. Epub 2013 Apr 4. PLoS Pathog. 2013. PMID: 23592994 Free PMC article.
-
Zn2+ blocks annealing of complementary single-stranded DNA in a sequence-selective manner.Sci Rep. 2014 Jun 26;4:5464. doi: 10.1038/srep05464. Sci Rep. 2014. PMID: 24965053 Free PMC article.
-
The CDC13-STN1-TEN1 complex stimulates Pol α activity by promoting RNA priming and primase-to-polymerase switch.Nat Commun. 2014 Dec 12;5:5762. doi: 10.1038/ncomms6762. Nat Commun. 2014. PMID: 25503194 Free PMC article.
-
Mechanism for priming DNA synthesis by yeast DNA polymerase α.Elife. 2013 Apr 2;2:e00482. doi: 10.7554/eLife.00482. Elife. 2013. PMID: 23599895 Free PMC article.
-
Mechanism of Concerted RNA-DNA Primer Synthesis by the Human Primosome.J Biol Chem. 2016 May 6;291(19):10006-20. doi: 10.1074/jbc.M116.717405. Epub 2016 Mar 14. J Biol Chem. 2016. PMID: 26975377 Free PMC article.
References
-
- Hamdan S. M., Richardson C. C. (2009) Motors, switches, and contacts in the replisome. Annu. Rev. Biochem. 78, 205–243 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
