

The human selenoprotein VCP-interacting membrane protein (VIMP) is non-globular and harbors a reductase function in an intrinsically disordered region
- PMID: 22700979
- PMCID: PMC3406722
- DOI: 10.1074/jbc.M112.346775
The human selenoprotein VCP-interacting membrane protein (VIMP) is non-globular and harbors a reductase function in an intrinsically disordered region
Abstract
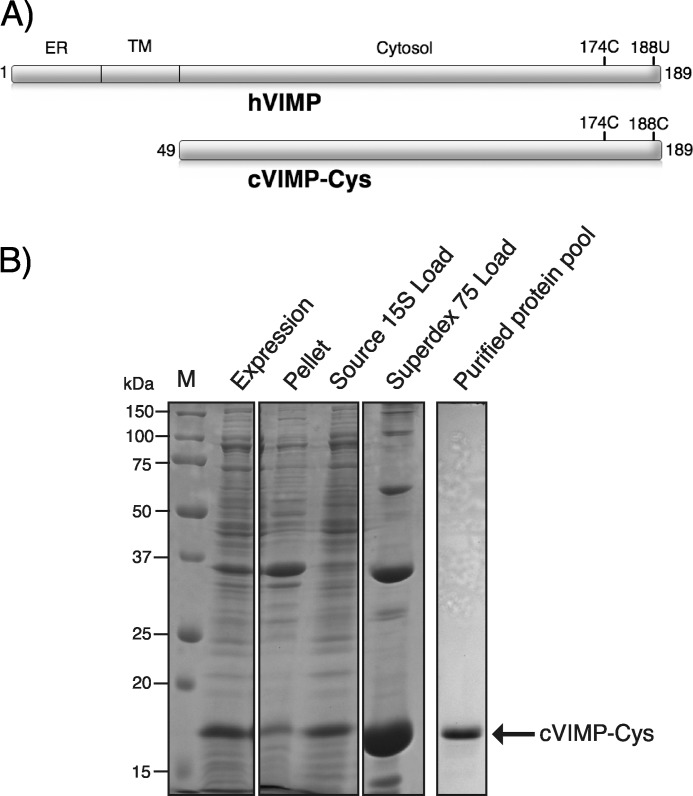
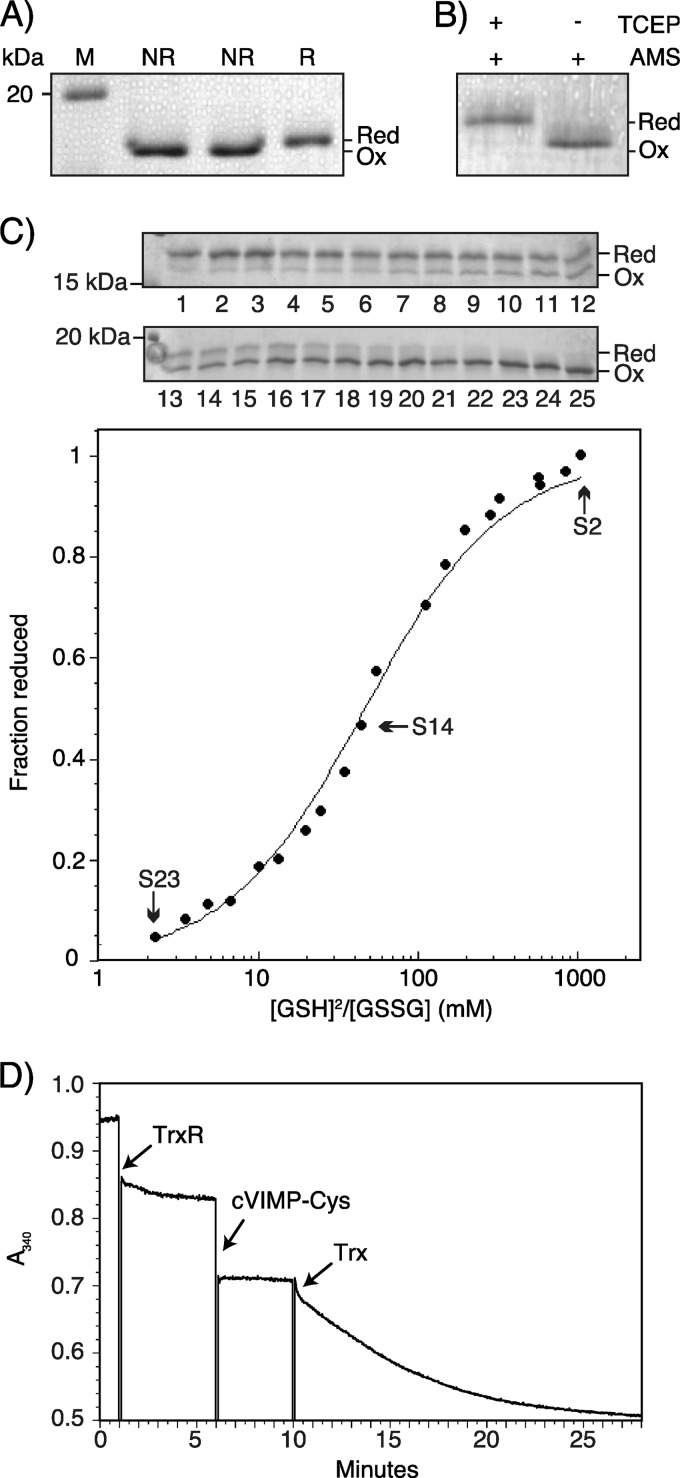
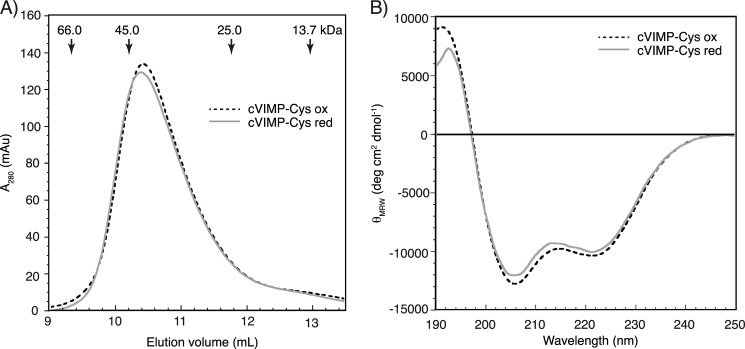
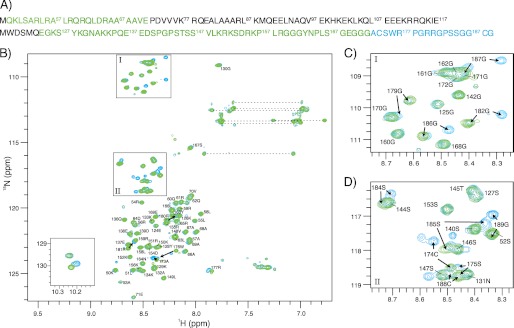
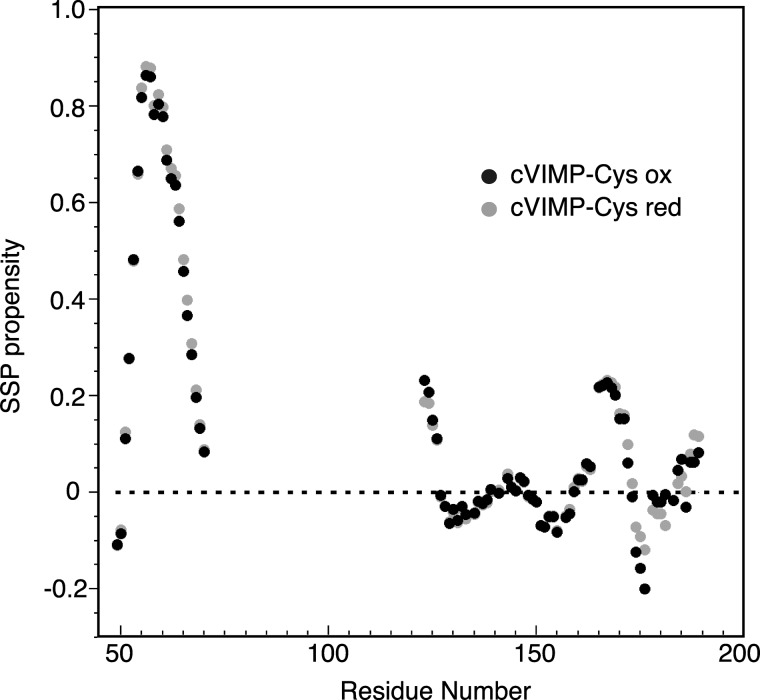
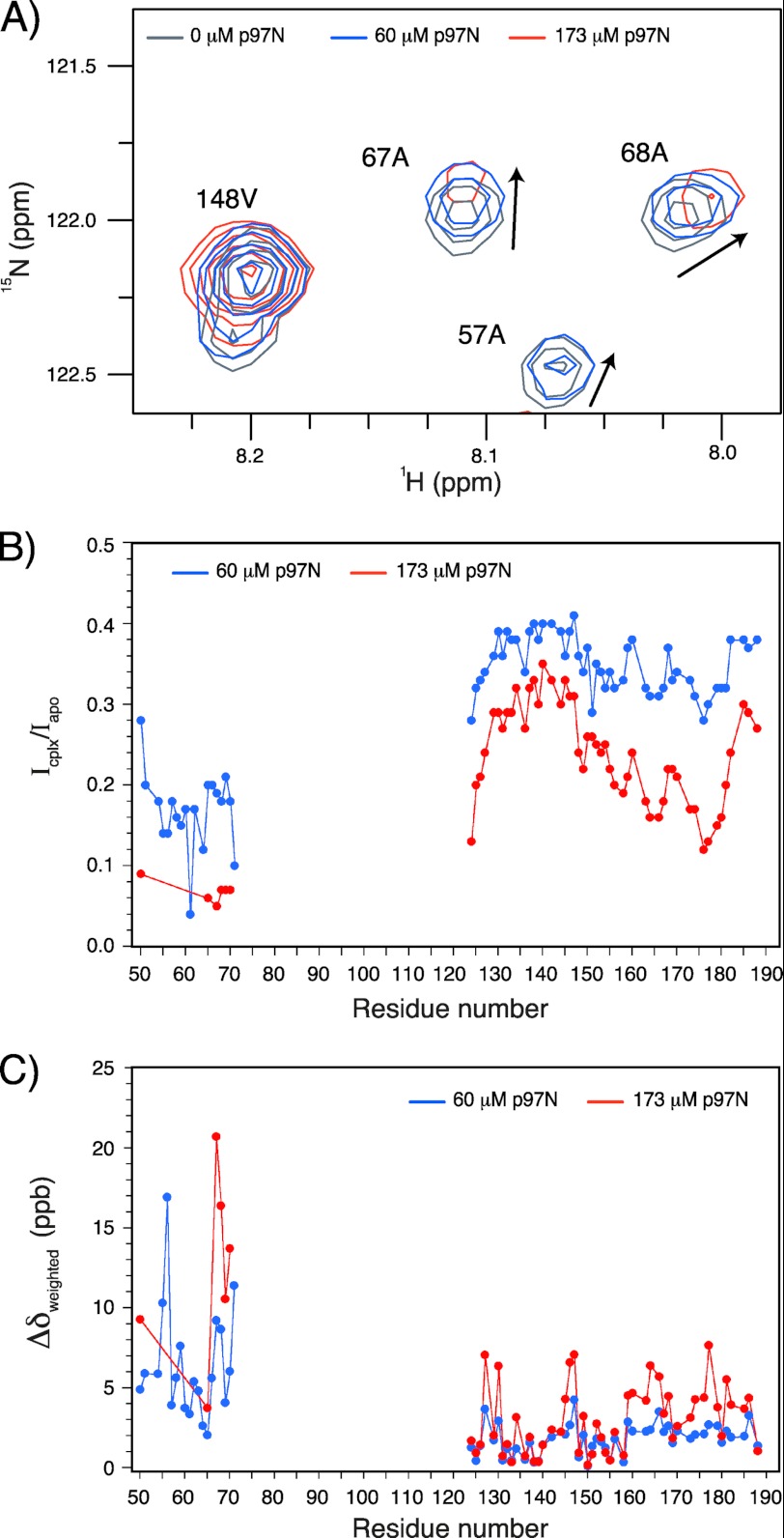
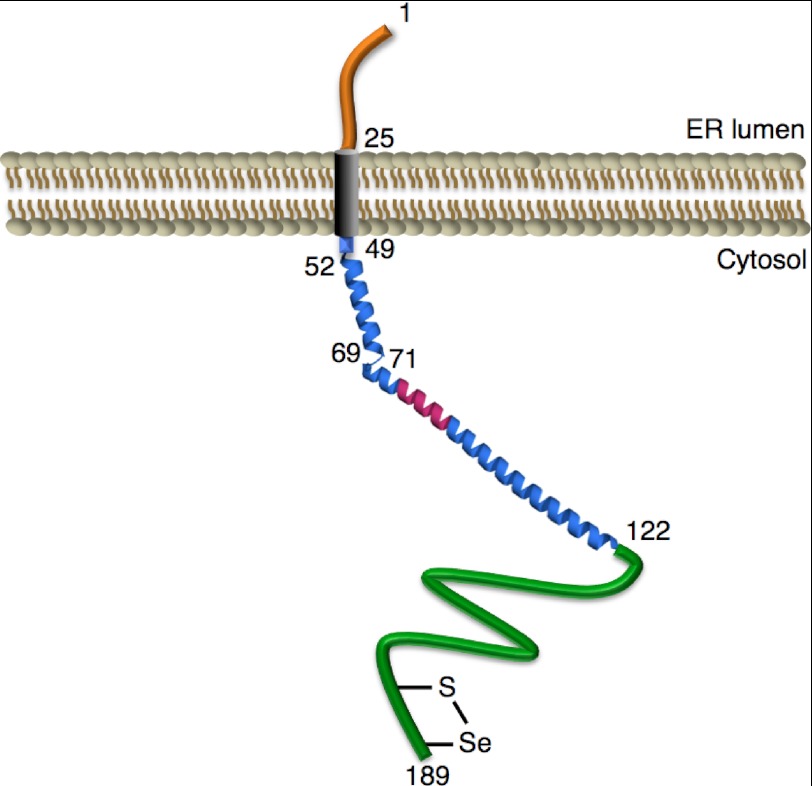
The human selenoprotein VIMP (VCP-interacting membrane protein)/SelS (selenoprotein S) localizes to the endoplasmic reticulum (ER) membrane and is involved in the process of ER-associated degradation (ERAD). To date, little is known about the presumed redox activity of VIMP, its structure and how these features might relate to the function of the protein in ERAD. Here, we use the recombinantly expressed cytosolic region of VIMP where the selenocysteine (Sec) in position 188 is replaced with a cysteine (a construct named cVIMP-Cys) to characterize redox and structural properties of the protein. We show that Cys-188 in cVIMP-Cys forms a disulfide bond with Cys-174, consistent with the presence of a Cys174-Sec188 selenosulfide bond in the native sequence. For the disulfide bond in cVIMP-Cys we determined the reduction potential to -200 mV, and showed it to be a good substrate of thioredoxin. Based on a biochemical and structural characterization of cVIMP-Cys using analytical gel filtration, CD and NMR spectroscopy in conjunction with bioinformatics, we propose a comprehensive overall structural model for the cytosolic region of VIMP. The data clearly indicate the N-terminal half to be comprised of two extended α-helices followed by a C-terminal region that is intrinsically disordered. Redox-dependent conformational changes in cVIMP-Cys were observed only in the vicinity of the two Cys residues. Overall, the redox properties observed for cVIMP-Cys are compatible with a function as a reductase, and we speculate that the plasticity of the intrinsically disordered C-terminal region allows the protein to access many different and structurally diverse substrates.
Figures
References
-
- Carvalho P., Goder V., Rapoport T. A. (2006) Distinct ubiquitin-ligase complexes define convergent pathways for the degradation of ER proteins. Cell 126, 361–373 - PubMed
-
- Denic V., Quan E. M., Weissman J. S. (2006) A luminal surveillance complex that selects misfolded glycoproteins for ER-associated degradation. Cell 126, 349–359 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
