

Lipid droplet formation on opposing sides of the endoplasmic reticulum
- PMID: 22701043
- PMCID: PMC3413221
- DOI: 10.1194/jlr.R028290
Lipid droplet formation on opposing sides of the endoplasmic reticulum
Abstract
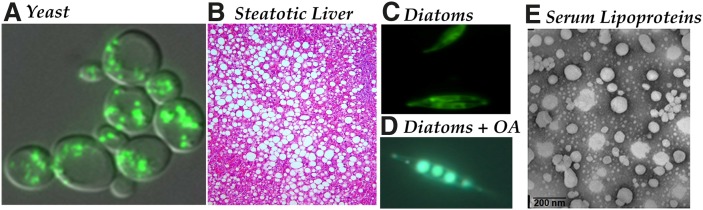
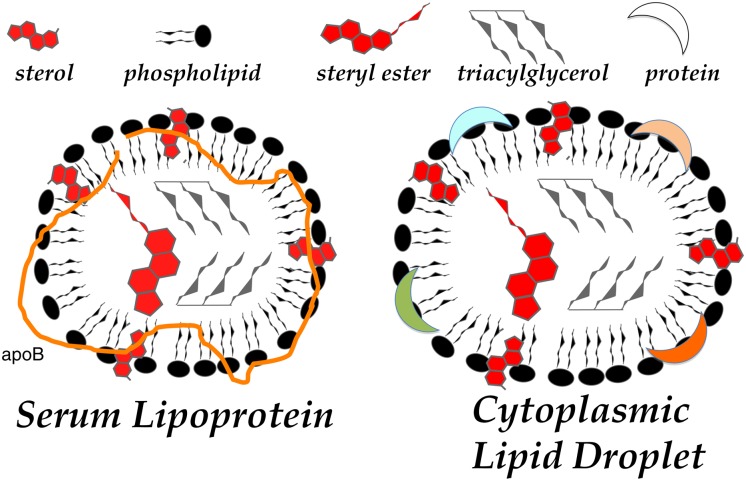
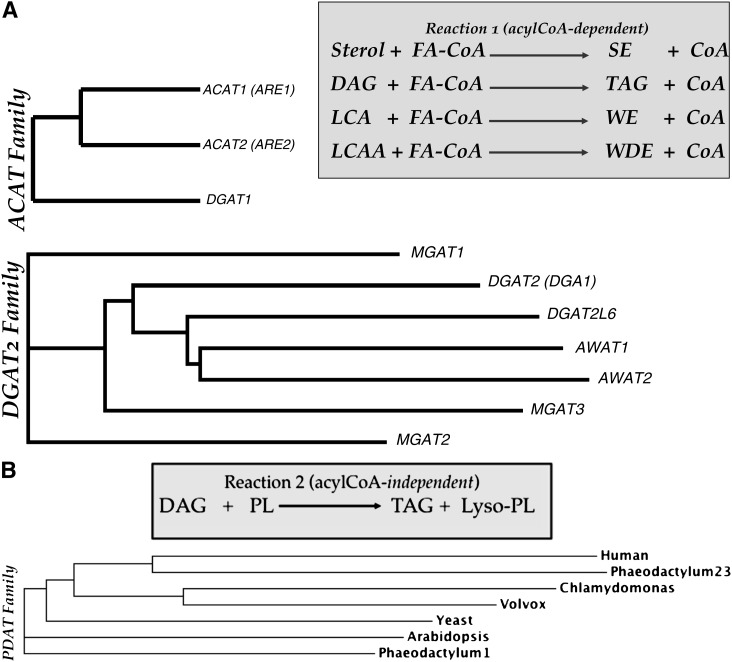
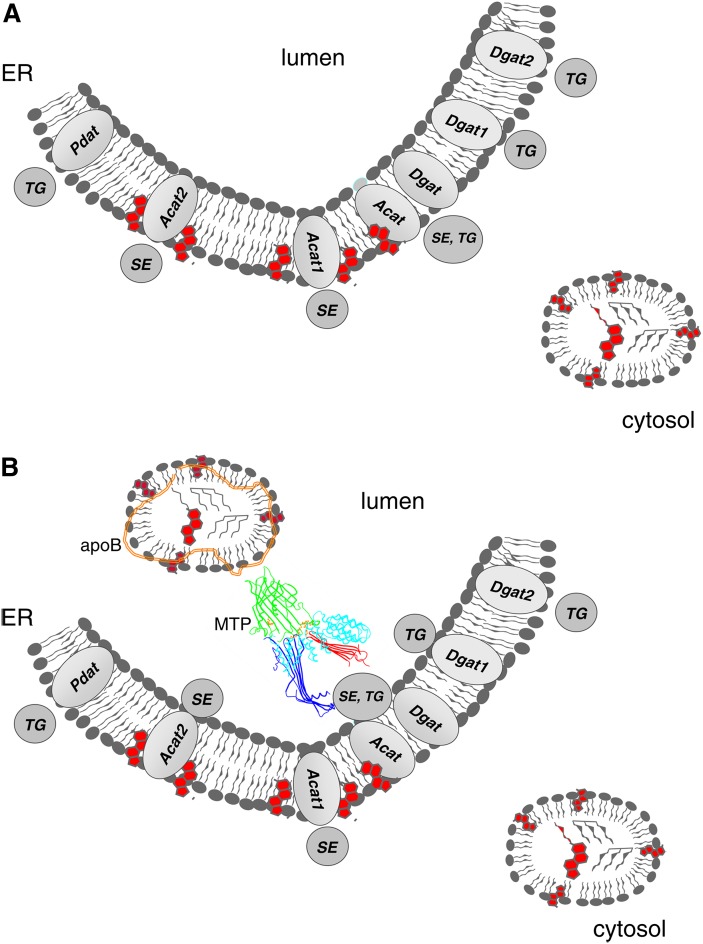
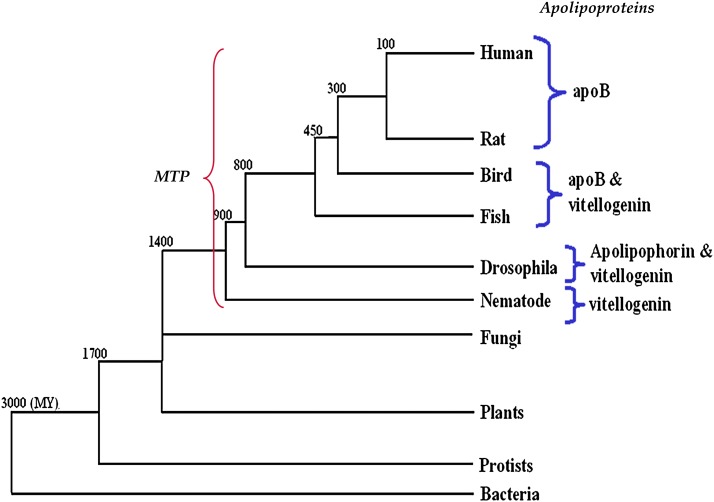
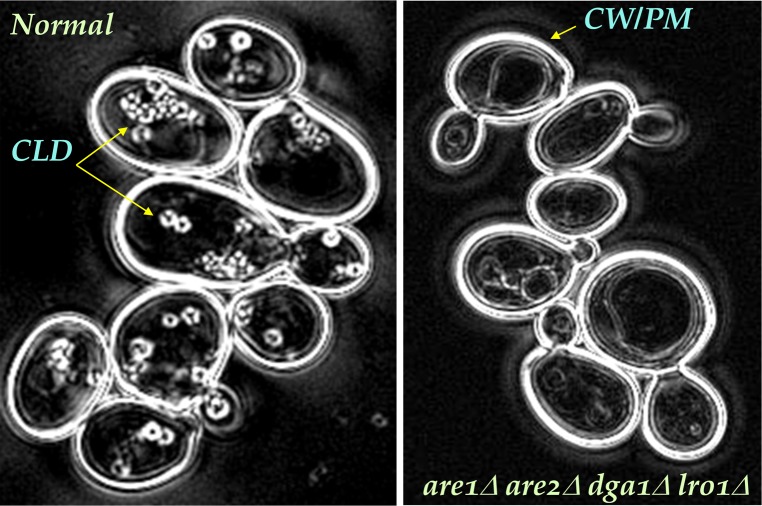
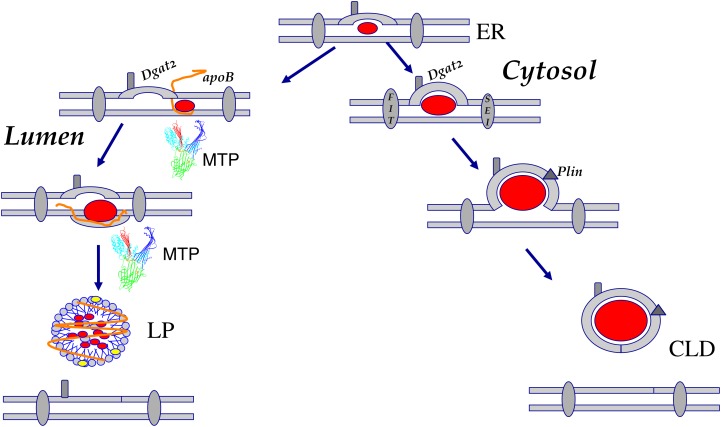
In animal cells, the primary repositories of esterified fatty acids and alcohols (neutral lipids) are lipid droplets that form on the lumenal and/or cytoplasmic side of the endoplasmic reticulum (ER) membrane. A monolayer of amphipathic lipids, intermeshed with key proteins, serves to solubilize neutral lipids as they are synthesized and desorbed. In specialized cells, mobilization of the lipid cargo for delivery to other tissues occurs by secretion of lipoproteins into the plasma compartment. Serum lipoprotein assembly requires an obligate structural protein anchor (apolipoprotein B) and a dedicated chaperone, microsomal triglyceride transfer protein. By contrast, lipid droplets that form on the cytoplasmic face of the ER lack an obligate protein scaffold or any required chaperone/lipid transfer protein. Mobilization of neutral lipids from the cytosol requires regulated hydrolysis followed by transfer of the products to different organelles or export from cells. Several proteins play a key role in controlling droplet number, stability, and catabolism; however, it is our premise that their formation initiates spontaneously, solely as a consequence of neutral lipid synthesis. This default pathway directs droplets into the cytoplasm where they accumulate in many lipid disorders.
Figures
References
-
- Wilson E. B. 1896. The Cell in Development and Inheritance Macmillan, New York.
-
- Chao F. F., Stiers D. L., Ontko J. A. 1986. Hepatocellular triglyceride synthesis and transfer to lipid droplets and nascent very low density lipoproteins. J. Lipid Res. 27: 1174–1181. - PubMed
-
- Turkish A., Sturley S. L. 2007. Regulation of triglyceride metabolism. I. Eukaryotic neutral lipid synthesis: “Many ways to skin ACAT or a DGAT”. Am. J. Physiol. Gastrointest. Liver Physiol. 292: G953–G957. - PubMed
-
- Stone S. J., Myers H. M., Watkins S. M., Brown B. E., Feingold K. R., Elias P. M., Farese R. V., Jr 2003. Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J. Biol. Chem. 279:11767–11176.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
