

Genetics Techniques for Thermococcus kodakarensis
- PMID: 22701112
- PMCID: PMC3370424
- DOI: 10.3389/fmicb.2012.00195
Genetics Techniques for Thermococcus kodakarensis
Abstract
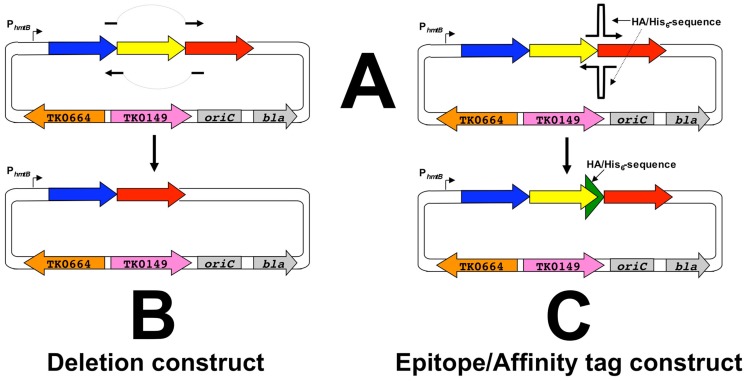
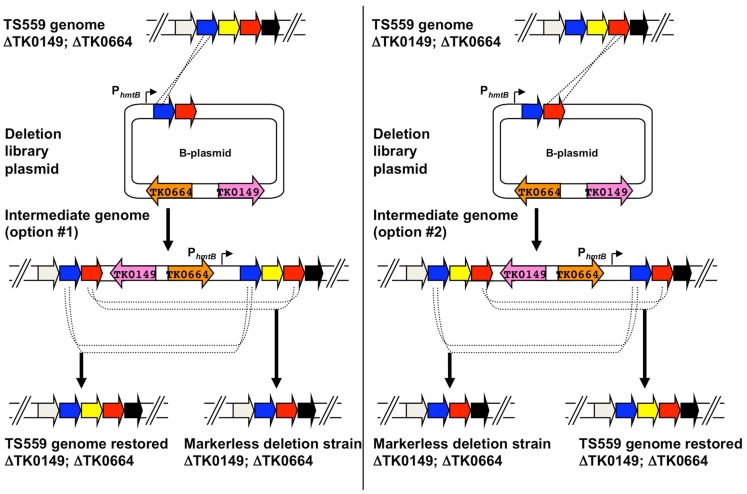
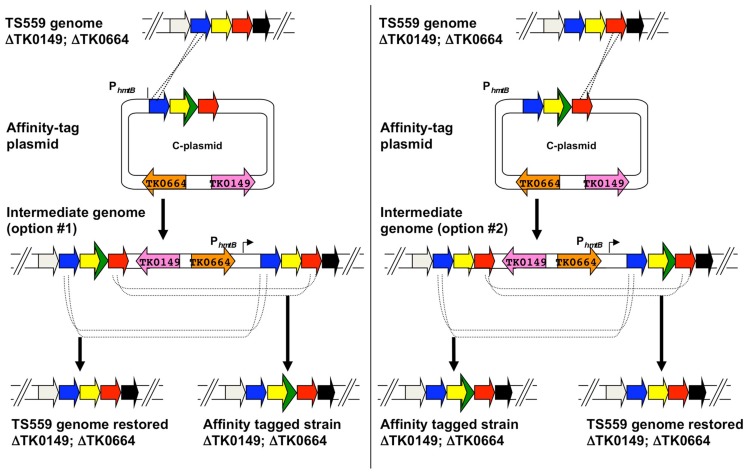
Thermococcus kodakarensis (T. kodakarensis) has emerged as a premier model system for studies of archaeal biochemistry, genetics, and hyperthermophily. This prominence is derived largely from the natural competence of T. kodakarensis and the comprehensive, rapid, and facile techniques available for manipulation of the T. kodakarensis genome. These genetic capacities are complemented by robust planktonic growth, simple selections, and screens, defined in vitro transcription and translation systems, replicative expression plasmids, in vivo reporter constructs, and an ever-expanding knowledge of the regulatory mechanisms underlying T. kodakarensis metabolism. Here we review the existing techniques for genetic and biochemical manipulation of T. kodakarensis. We also introduce a universal platform to generate the first comprehensive deletion and epitope/affinity tagged archaeal strain libraries.
Keywords: Thermococcales; archaea; genetics; hyperthermophilic; recombination; transcription.
Figures
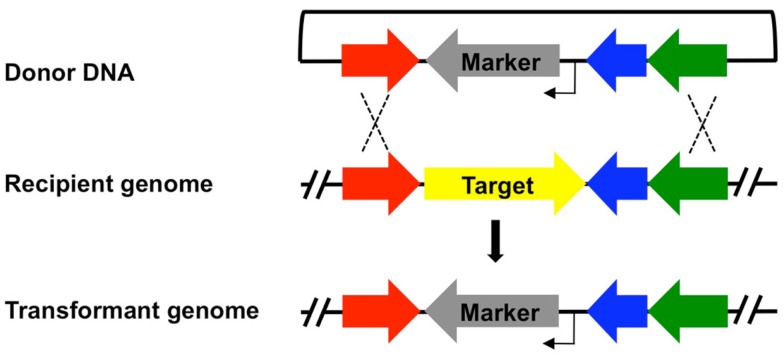
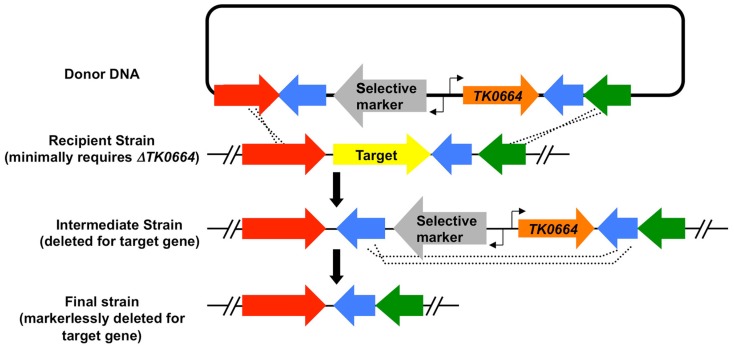
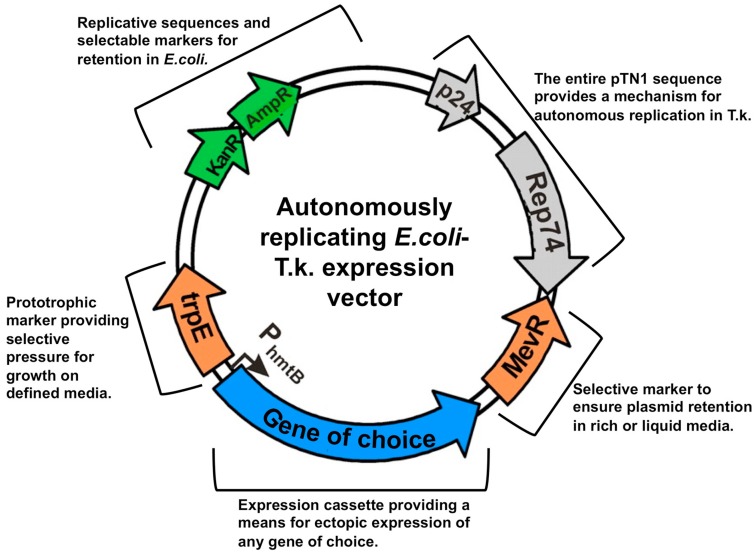
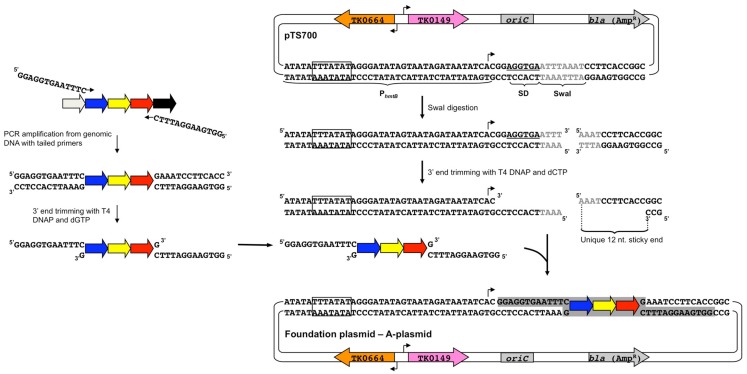
References
Grants and funding
LinkOut - more resources
Full Text Sources