

Interspecific nematode signals regulate dispersal behavior
- PMID: 22701701
- PMCID: PMC3368880
- DOI: 10.1371/journal.pone.0038735
Interspecific nematode signals regulate dispersal behavior
Abstract
Background: Dispersal is an important nematode behavior. Upon crowding or food depletion, the free living bacteriovorus nematode Caenorhabditis elegans produces stress resistant dispersal larvae, called dauer, which are analogous to second stage juveniles (J2) of plant parasitic Meloidogyne spp. and infective juveniles (IJ)s of entomopathogenic nematodes (EPN), e.g., Steinernema feltiae. Regulation of dispersal behavior has not been thoroughly investigated for C. elegans or any other nematode species. Based on the fact that ascarosides regulate entry in dauer stage as well as multiple behaviors in C. elegans adults including mating, avoidance and aggregation, we hypothesized that ascarosides might also be involved in regulation of dispersal behavior in C. elegans and for other nematodes such as IJ of phylogenetically related EPNs.
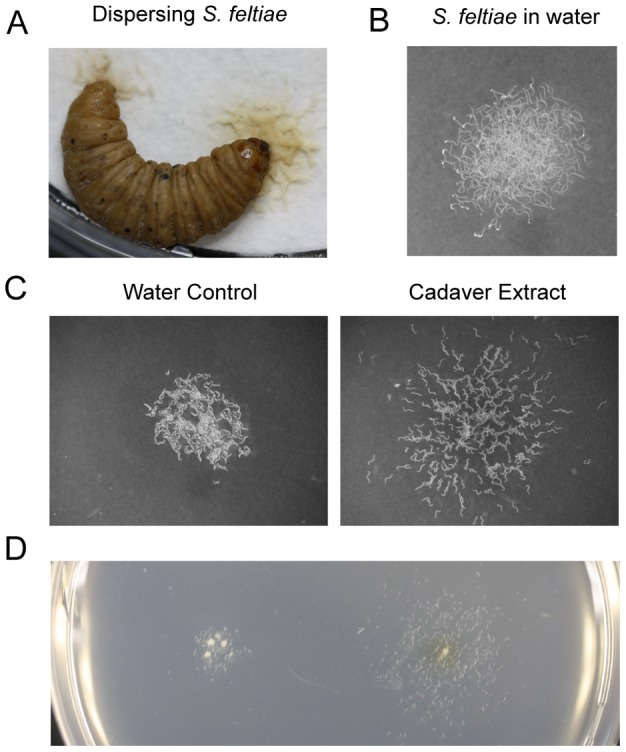
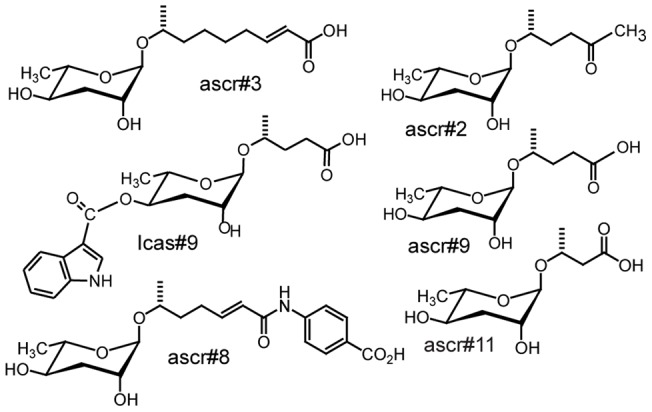
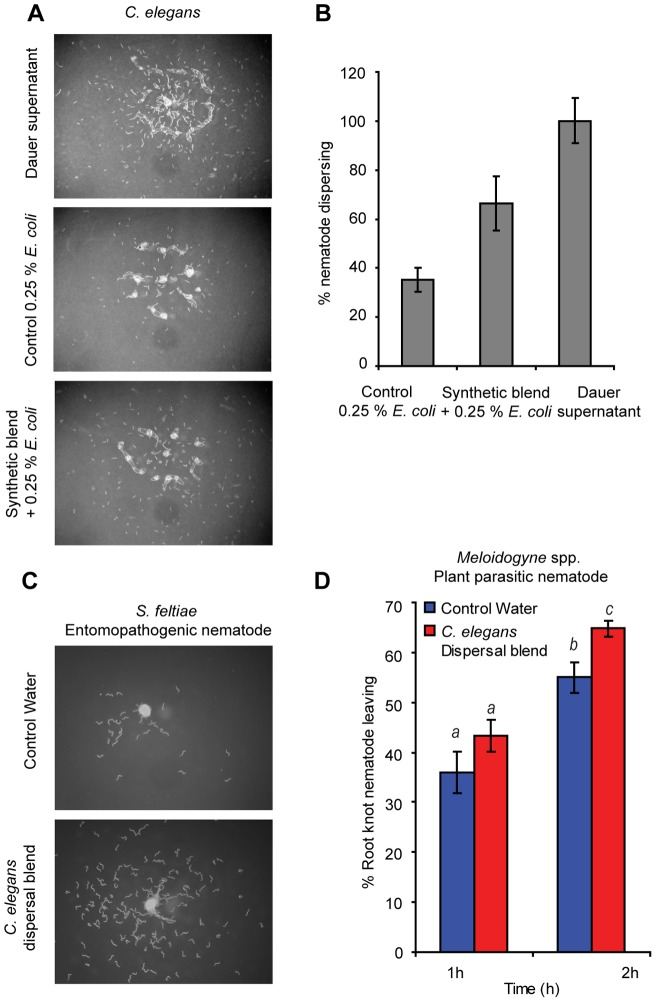
Methodology/principal findings: Liquid chromatography-mass spectrometry analysis of C. elegans dauer conditioned media, which shows strong dispersing activity, revealed four known ascarosides (ascr#2, ascr#3, ascr#8, icas#9). A synthetic blend of these ascarosides at physiologically relevant concentrations dispersed C. elegans dauer in the presence of food and also caused dispersion of IJs of S. feltiae and J2s of plant parasitic Meloidogyne spp. Assay guided fractionation revealed structural analogs as major active components of the S. feltiae (ascr#9) and C. elegans (ascr#2) dispersal blends. Further analysis revealed ascr#9 in all Steinernema spp. and Heterorhabditis spp. infected insect host cadavers.
Conclusions/significance: Ascaroside blends represent evolutionarily conserved, fundamentally important communication systems for nematodes from diverse habitats, and thus may provide sustainable means for control of parasitic nematodes.
Conflict of interest statement
Figures

References
-
- Kaya HK, Gaugler R. Entomopathogenic Nematodes. Annu Rev Entomol. 1993;38:181–206.
-
- Barriere A, Felix MA. High local genetic diversity and low outcrossing rate in Caenorhabditis elegans natural populations. Curr Biol. 2005;15:1176–1184. - PubMed
-
- Golden JW, Riddle DL. The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev Biol. 1984;102:368–378. - PubMed
-
- Golden JW, Riddle DL. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science. 1982;218:578–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
