

Deviation of innate circadian period from 24 h reduces longevity in mice
- PMID: 22702406
- PMCID: PMC3526942
- DOI: 10.1111/j.1474-9726.2012.00846.x
Deviation of innate circadian period from 24 h reduces longevity in mice
Abstract
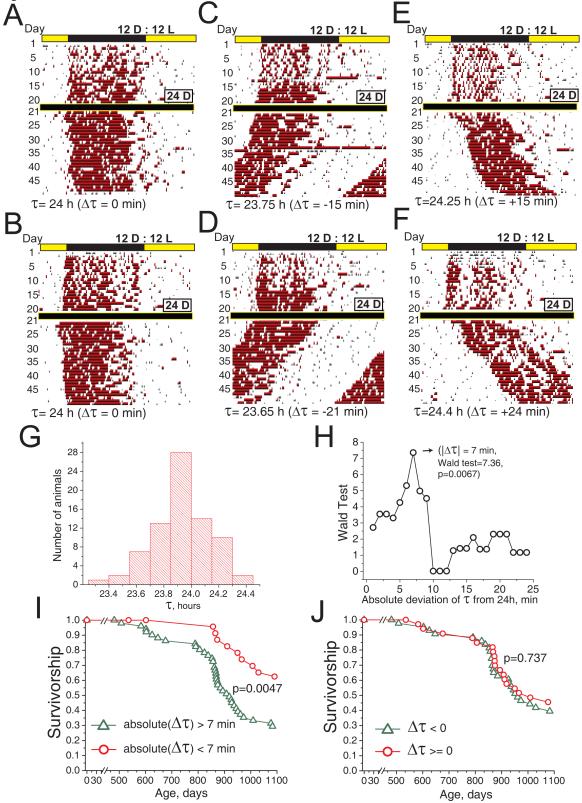
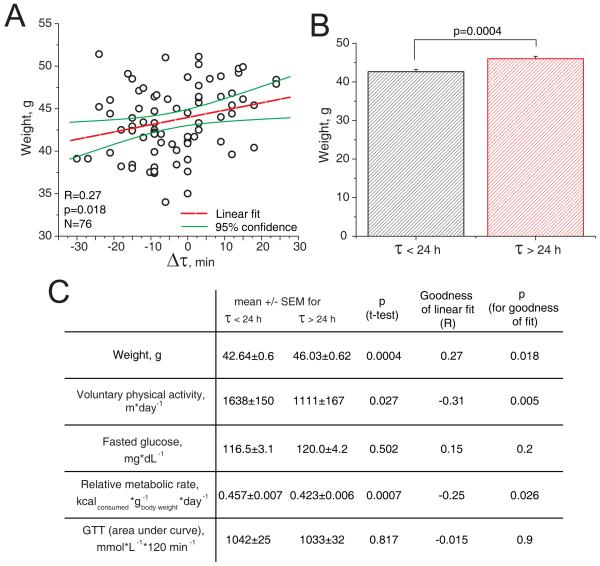
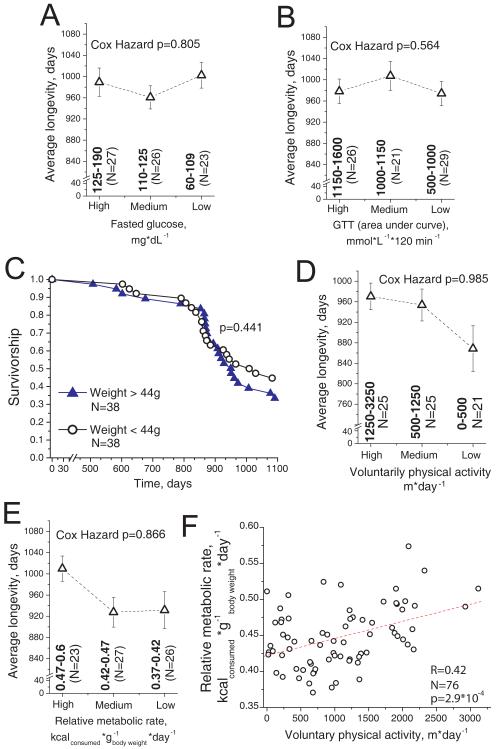
The variation of individual life spans, even in highly inbred cohorts of animals and under strictly controlled environmental conditions, is substantial and not well understood. This variation in part could be due to epigenetic variation, which later affects the animal's physiology and ultimately longevity. Identification of the physiological properties that impact health and life span is crucial for longevity research and the development of anti-aging therapies. Here, we measured individual circadian and metabolic characteristics in a cohort of inbred F1 hybrid mice and correlated these parameters to their life spans. We found that mice with innate circadian periods close to 24 h (revealed during 30 days of housing in total darkness) enjoyed nearly 20% longer life spans than their littermates, which had shorter or longer innate circadian periods. These findings show that maintenance of a 24-h intrinsic circadian period is a positive predictor of longevity. Our data suggest that circadian period may be used to predict individual longevity and that processes that control innate circadian period affect aging.
© 2012 The Authors. Aging Cell © 2012 Blackwell Publishing Ltd/Anatomical Society of Great Britain and Ireland.
Figures
Similar articles
-
Circadian rhythms of body temperature and locomotor activity in aging BALB/c mice: early and late life span predictors.Biogerontology. 2016 Aug;17(4):703-14. doi: 10.1007/s10522-016-9635-y. Epub 2016 Jan 28. Biogerontology. 2016. PMID: 26820297
-
Circadian rhythms, aging, and life span in mammals.Physiology (Bethesda). 2011 Aug;26(4):225-35. doi: 10.1152/physiol.00012.2011. Physiology (Bethesda). 2011. PMID: 21841071 Review.
-
Effect of feeding regimens on circadian rhythms: implications for aging and longevity.Aging (Albany NY). 2010 Dec 11;2(1):7-27. doi: 10.18632/aging.100116. Aging (Albany NY). 2010. PMID: 20228939 Free PMC article. Review.
-
Sleep-length differences are associated with altered longevity in the fruit fly Drosophila melanogaster.Biol Open. 2020 Sep 16;9(9):bio054361. doi: 10.1242/bio.054361. Biol Open. 2020. PMID: 32938639 Free PMC article.
-
The aging clock: circadian rhythms and later life.J Clin Invest. 2017 Feb 1;127(2):437-446. doi: 10.1172/JCI90328. Epub 2017 Feb 1. J Clin Invest. 2017. PMID: 28145903 Free PMC article. Review.
Cited by
-
Epigenetic regulation of aging: implications for interventions of aging and diseases.Signal Transduct Target Ther. 2022 Nov 7;7(1):374. doi: 10.1038/s41392-022-01211-8. Signal Transduct Target Ther. 2022. PMID: 36336680 Free PMC article. Review.
-
Systemic regulation of mammalian ageing and longevity by brain sirtuins.Nat Commun. 2014 Jun 26;5:4211. doi: 10.1038/ncomms5211. Nat Commun. 2014. PMID: 24967620 Free PMC article. Review.
-
The brain, sirtuins, and ageing.Nat Rev Neurosci. 2017 May 18;18(6):362-374. doi: 10.1038/nrn.2017.42. Nat Rev Neurosci. 2017. PMID: 28515492 Review.
-
Calorie restriction and sirtuins revisited.Genes Dev. 2013 Oct 1;27(19):2072-85. doi: 10.1101/gad.227439.113. Genes Dev. 2013. PMID: 24115767 Free PMC article. Review.
-
The intricate link between circadian rhythms and aging: can resetting our circadian clock hold the key to longevity?Biogerontology. 2025 Aug 6;26(4):156. doi: 10.1007/s10522-025-10299-8. Biogerontology. 2025. PMID: 40770521 Review.
References
-
- Andrikopoulos S, Blair AR, Deluca N, Fam BC, Proietto J. Evaluating the glucose tolerance test in mice. Am J Physiol Endocrinol Metab. 2008;295:E1323–1332. - PubMed
-
- Anisimov VN, Zavarzina NY, Zabezhinski MA, Popovich IG, Zimina OA, Shtylick AV, Arutjunyan AV, Oparina TI, Prokopenko VM, Mikhalski AI, et al. Melatonin increases both life span and tumor incidence in female CBA mice. J Gerontol A Biol Sci Med Sci. 2001;56:B311–323. - PubMed
-
- Bartke A. Delayed aging in Ames dwarf mice. Relationships to endocrine function and body size. Results Probl Cell Differ. 2000;29:181–202. - PubMed
