

Toll-like receptor 7 is required for effective adaptive immune responses that prevent persistent virus infection
- PMID: 22704624
- PMCID: PMC3377981
- DOI: 10.1016/j.chom.2012.04.016
Toll-like receptor 7 is required for effective adaptive immune responses that prevent persistent virus infection
Abstract
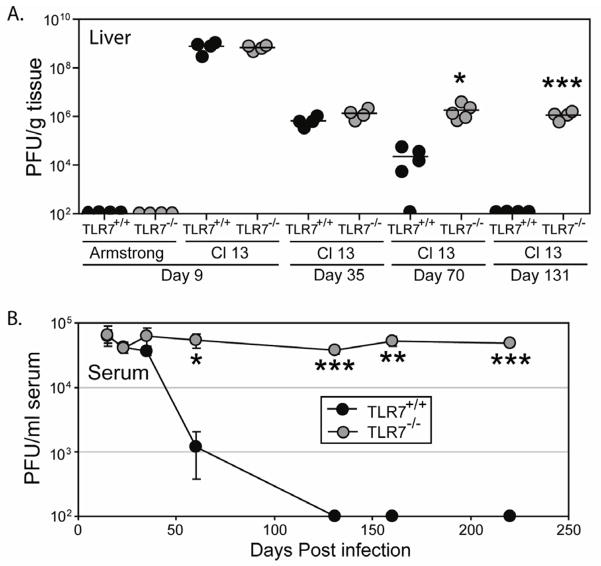
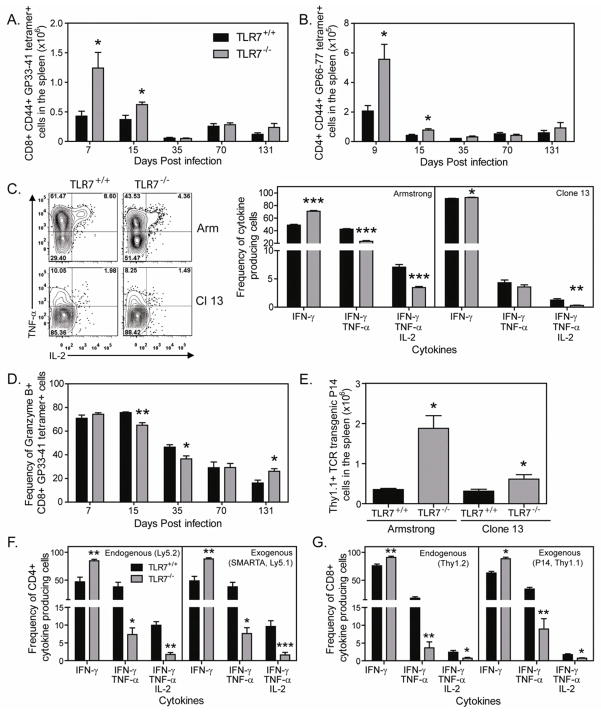
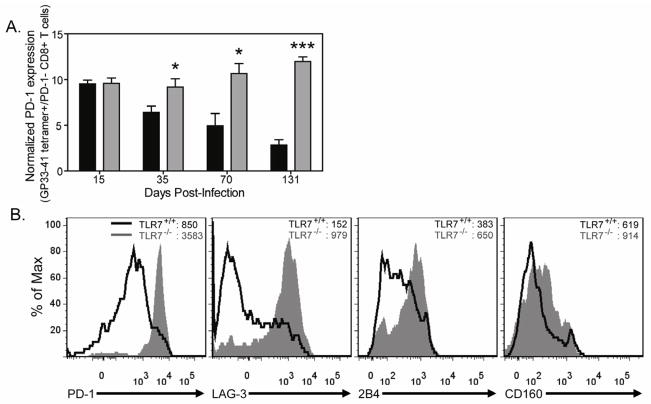
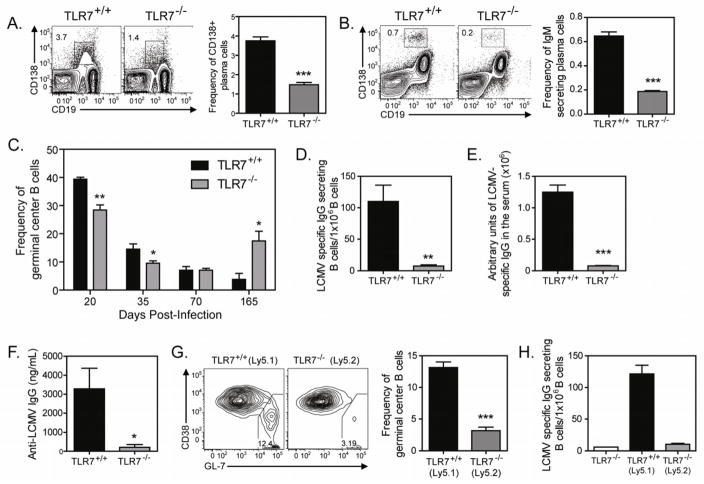
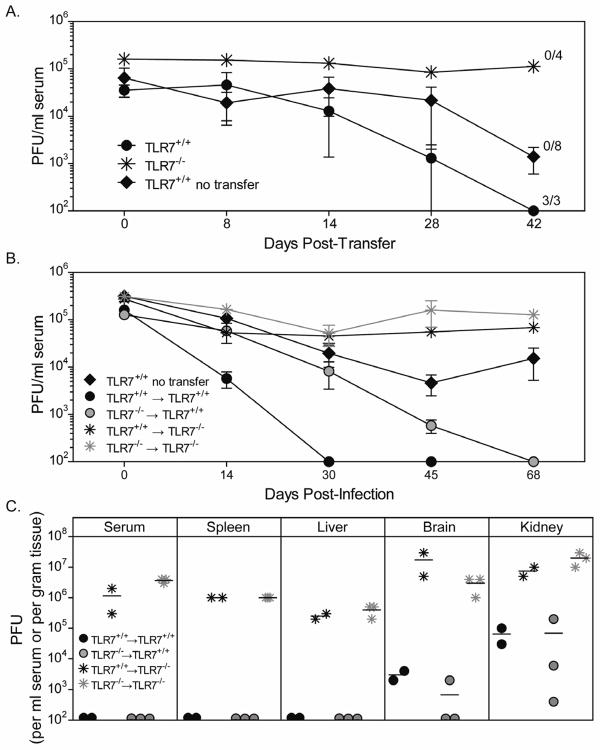
TLR7 is an innate signaling receptor that recognizes single-stranded viral RNA and is activated by viruses that cause persistent infections. We show that TLR7 signaling dictates either clearance or establishment of life-long chronic infection by lymphocytic choriomeningitis virus (LCMV) Cl 13 but does not affect clearance of the acute LCMV Armstrong 53b strain. TLR7(-/-) mice infected with LCMV Cl 13 remained viremic throughout life from defects in the adaptive antiviral immune response-notably, diminished T cell function, exacerbated T cell exhaustion, decreased plasma cell maturation, and negligible antiviral antibody production. Adoptive transfer of TLR7(+/+) LCMV immune memory cells that enhanced clearance of persistent LCMV Cl 13 infection in TLR7(+/+) mice failed to purge LCMV Cl 13 infection in TLR7(-/-) mice, demonstrating that a TLR7-deficient environment renders antiviral responses ineffective. Therefore, methods that promote TLR7 signaling are promising treatment strategies for chronic viral infections.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Lymphocyte choriomeningitis virus plays hide-and-seek with type 1 interferon.Cell Host Microbe. 2012 Jun 14;11(6):553-5. doi: 10.1016/j.chom.2012.05.007. Cell Host Microbe. 2012. PMID: 22704613 Review.
References
-
- Ambach A, Bonnekoh B, Nguyen M, Schon MP, Gollnick H. Imiquimod, a Toll-like receptor-7 agonist, induces perforin in cytotoxic T lymphocytes in vitro. Mol Immunol. 2004;40:1307–1314. - PubMed
-
- Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. - PubMed
-
- Berger DP, Homann D, Oldstone MB. Defining parameters for successful immunocytotherapy of persistent viral infection. Virology. 2000;266:257–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
