

The emergence and maintenance of sickle cell hotspots in the Mediterranean
- PMID: 22704979
- PMCID: PMC3438445
- DOI: 10.1016/j.meegid.2012.06.001
The emergence and maintenance of sickle cell hotspots in the Mediterranean
Abstract
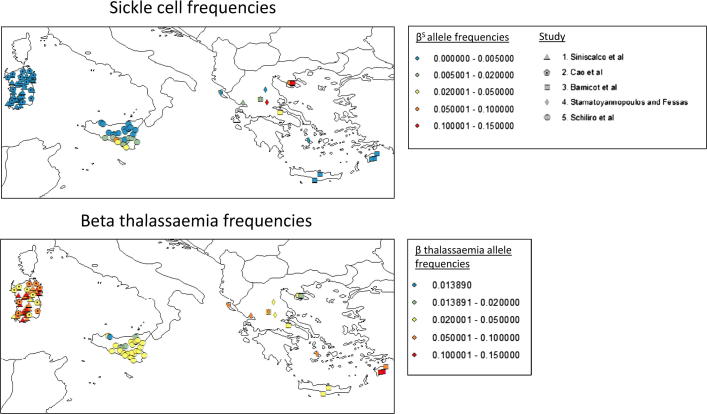
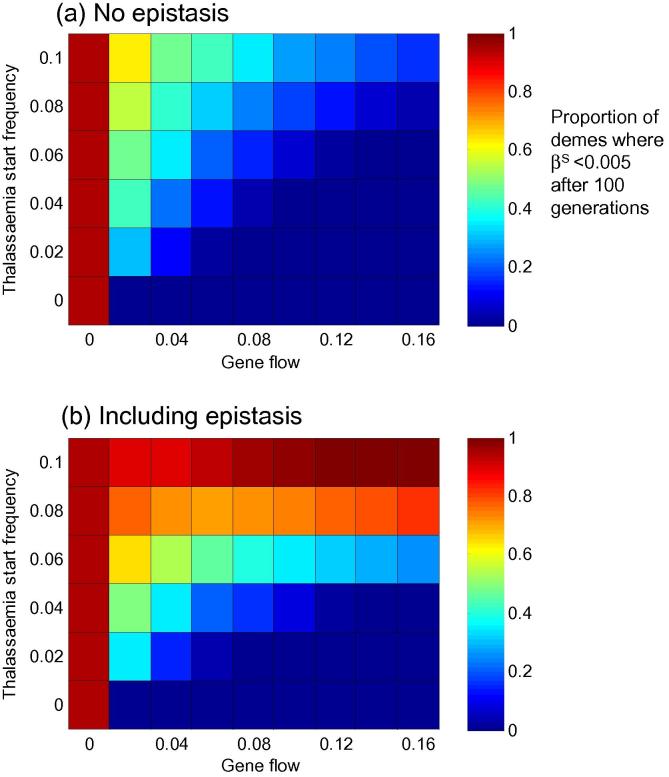
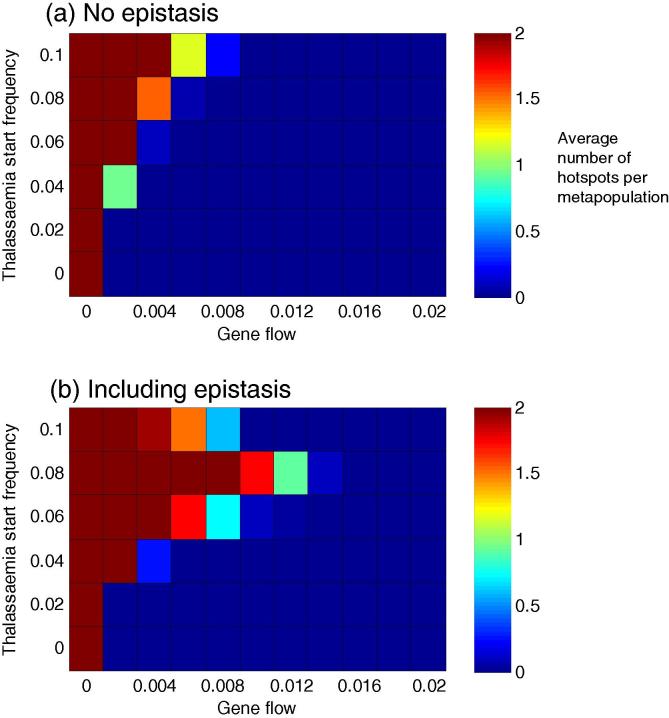
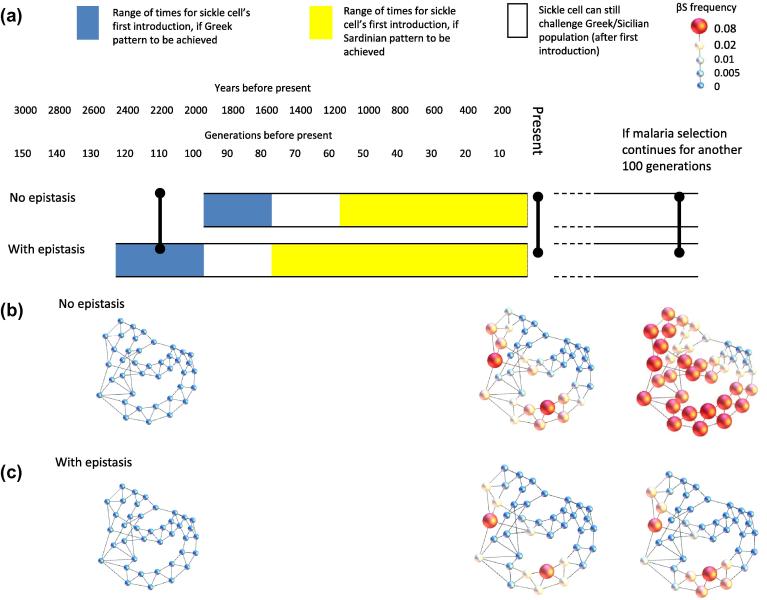
Genetic disorders of haemoglobin (haemoglobinopathies), including the thalassaemias and sickle cell anaemia, abound in historically malarious regions, due to the protection they provide against death from severe malaria. Despite the overall spatial correlation between malaria and these disorders, inter-population differences exist in the precise combinations of haemoglobinopathies observed. Greece and Italy present a particularly interesting case study: their high frequencies of beta thalassaemia speak to a history of intense malaria selection, yet they possess very little of the strongly malaria protective mutation responsible for sickle cell anaemia, despite historical migrational links with Africa where high frequencies of sickle cell occur. Twentieth century surveys of beta thalassaemia and sickle cell in Greece, Sicily and Sardinia have revealed striking sickle cell 'hotspots' - places where the frequency of sickle cell approaches that seen in Africa while neighbouring populations remain relatively sickle cell free. It remains unclear how these hotspots have been maintained over time without sickle cell spreading throughout the region. Here we use a metapopulation model to show that (i) epistasis between the alpha and beta forms of thalassaemia can restrict the spread of sickle cell through a network of linked subpopulations and (ii) the emergence of sickle cell hotspots requires relatively low levels of gene flow, but the aforementioned epistasis increases the chances of hotspots forming.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Adams J.W., Kasakoff A.B. Factors Underlying Endogamous Group Size. In: Smith C., editor. Regional Analysis. Academic Press; London: 1976. p. 149.
-
- Barnicot N.A., Allison A.C., Blumberg B.S., Deliyannis G., Krimbas C., Ballas A. Haemoglobin types in Greek populations. Ann. Hum. Genet. 1963;26:229–236. - PubMed
-
- Bastian, M., Heymann, S., Jacomy, M., 2009. Gephi: An Open Source Software for Exploring and Manipulating Networks.
-
- Boussiou M., Loukopoulos D., Christakis J., Fessas P. The origin of the sickle mutation in Greece; evidence from β(S) globin gene cluster polymorphisms. Hemoglobin. 1991;15:459–467. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
