

Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response
- PMID: 22705106
- PMCID: PMC3412146
- DOI: 10.1016/j.immuni.2012.03.022
Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response
Abstract
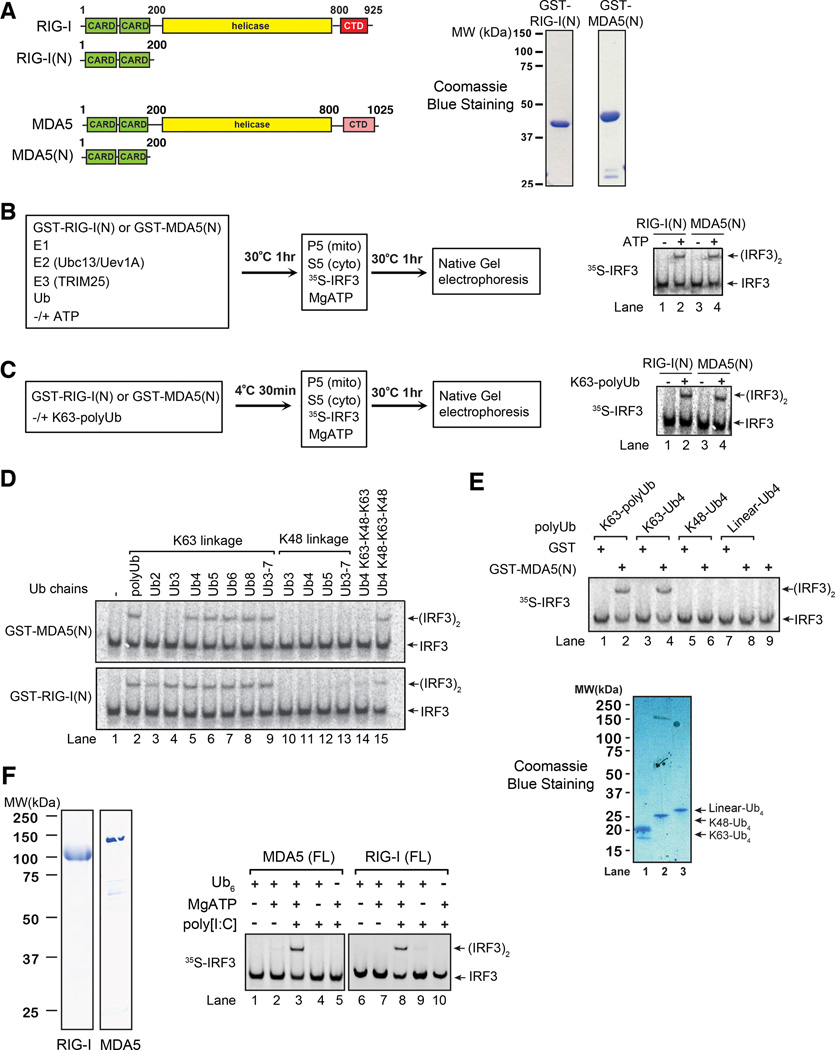
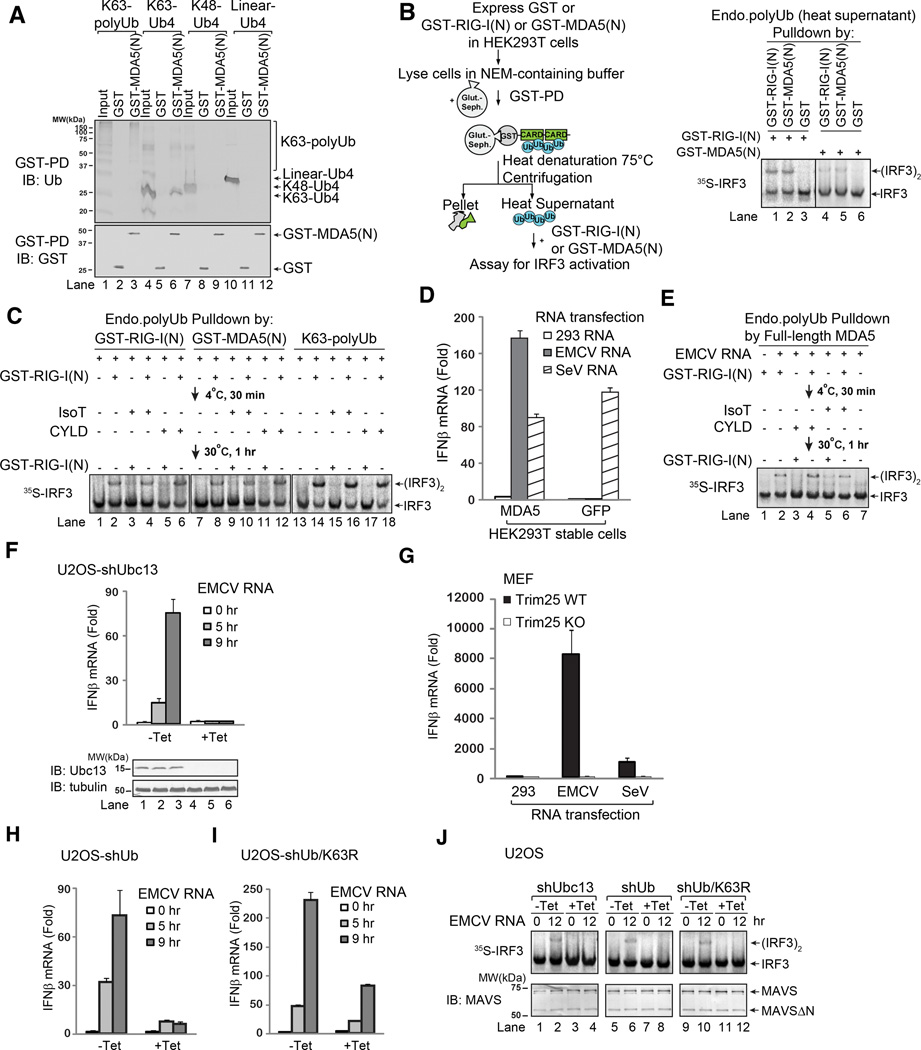
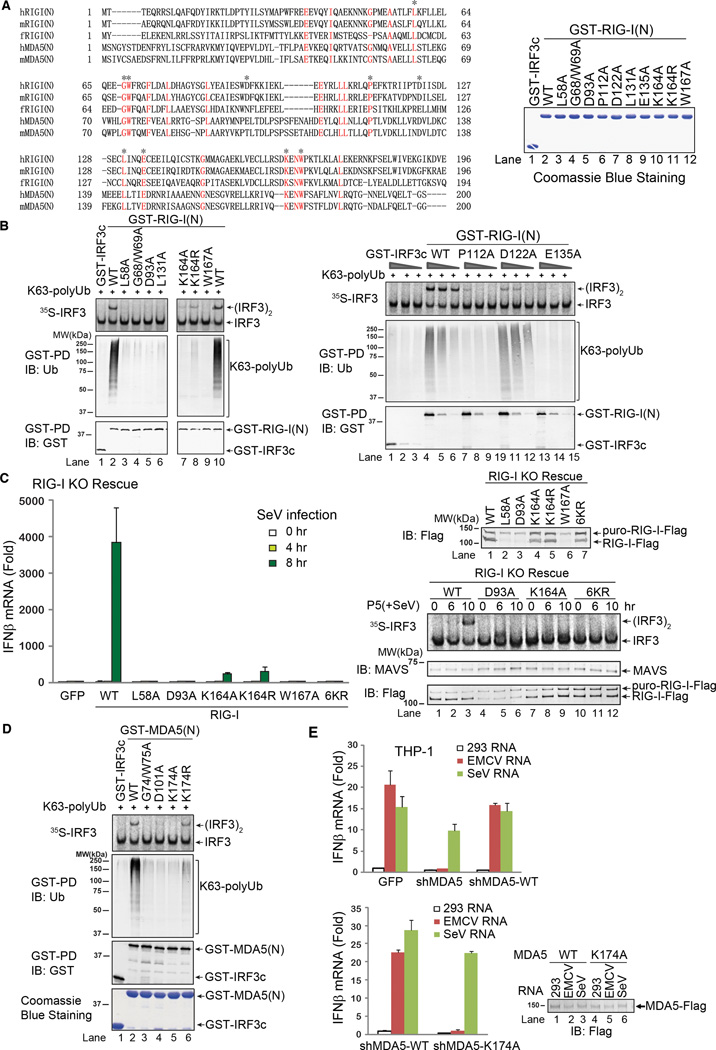
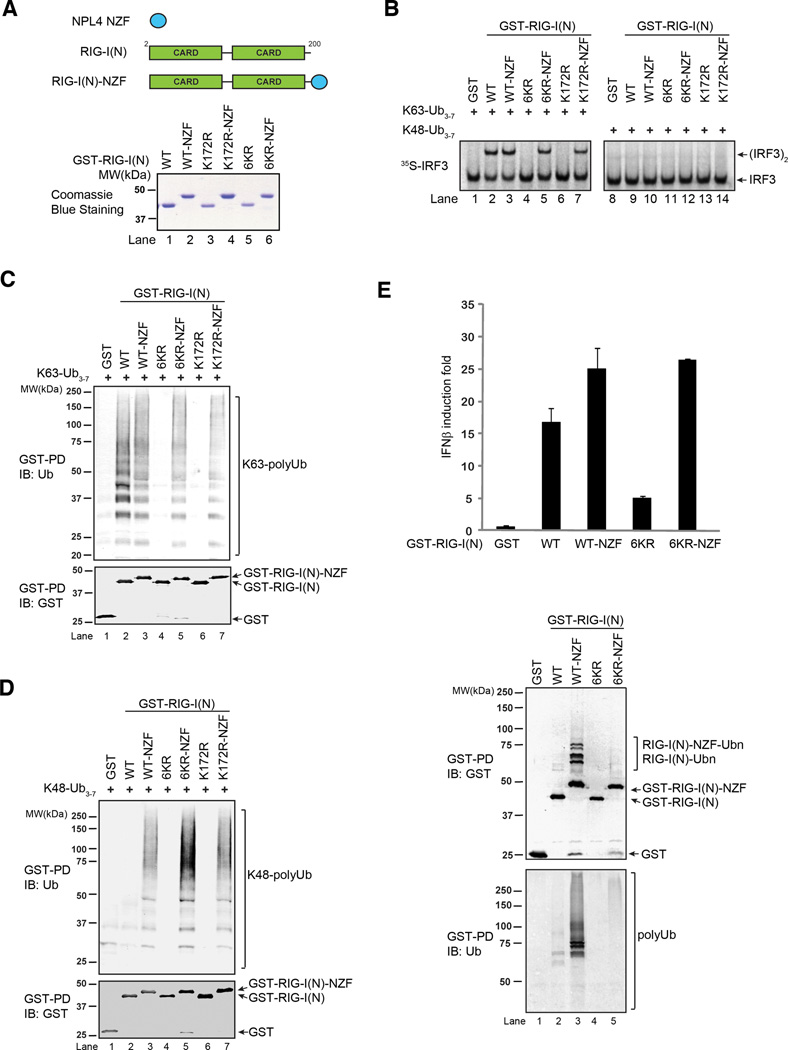
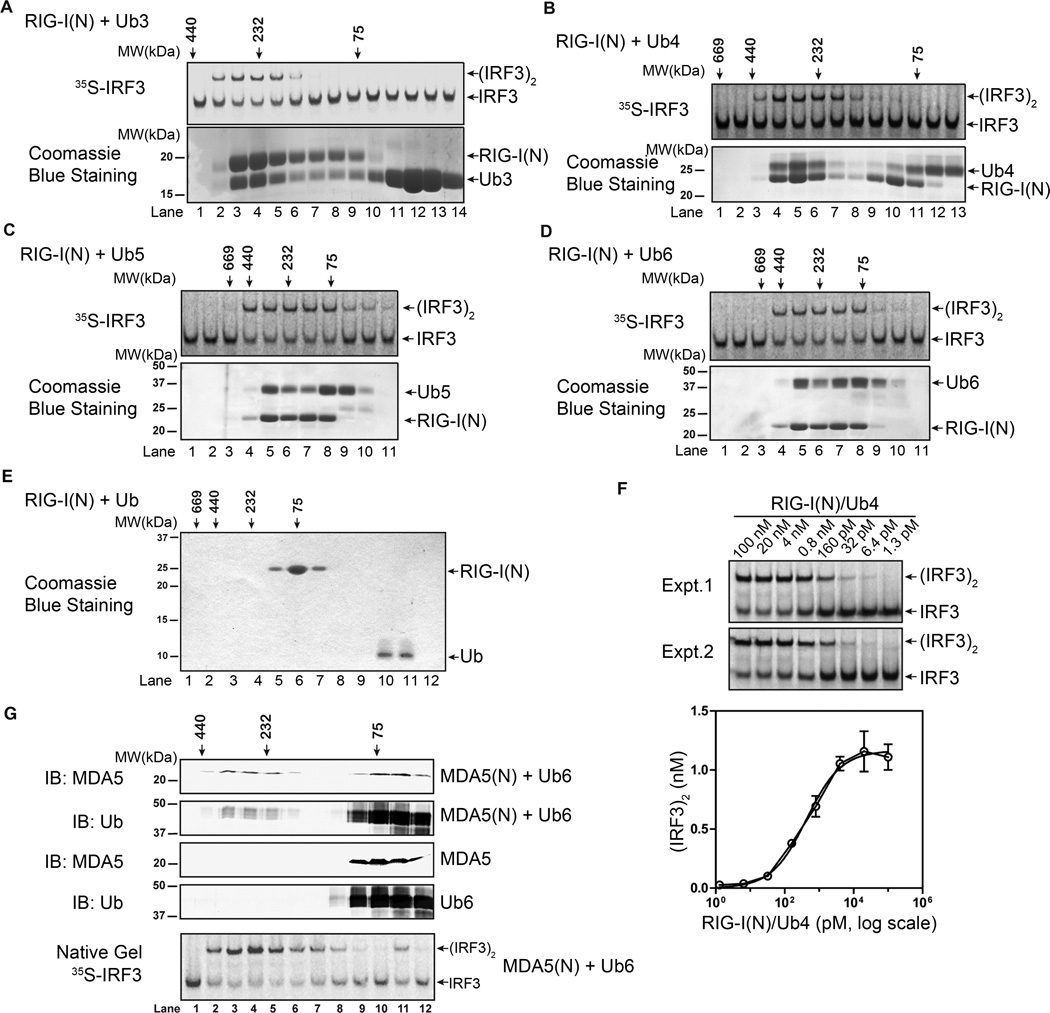
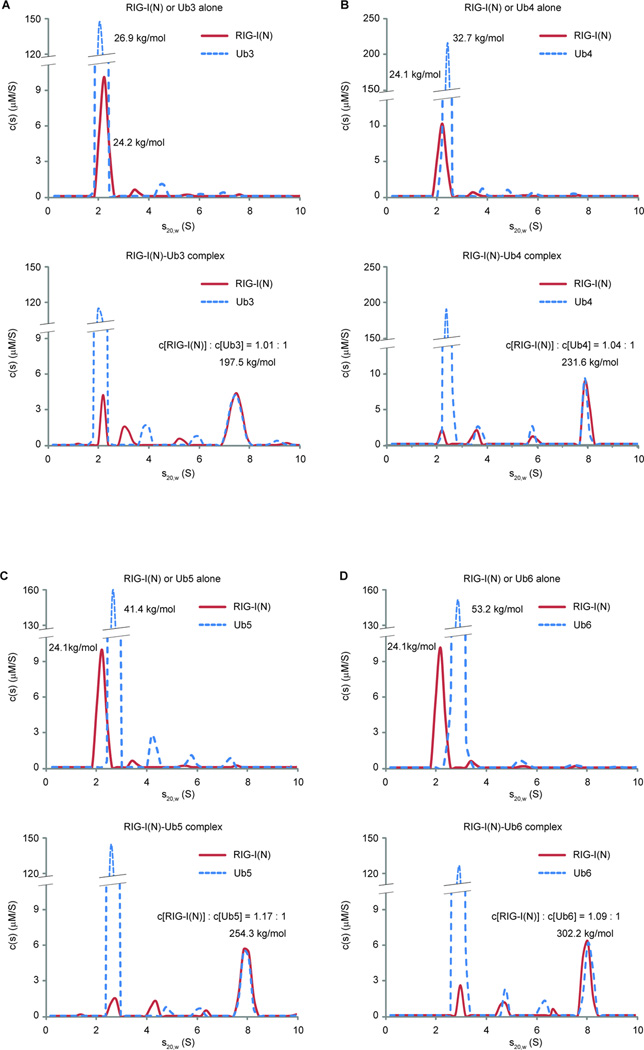
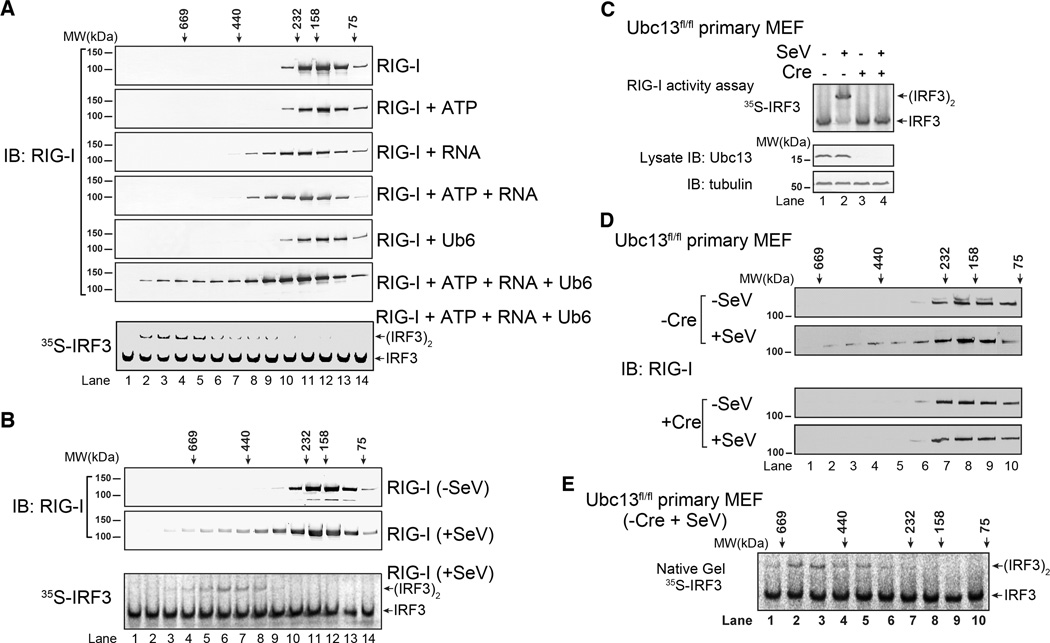
RIG-I and MDA5 detect viral RNA in the cytoplasm and activate signaling cascades leading to the production of type-I interferons. RIG-I is activated through sequential binding of viral RNA and unanchored lysine-63 (K63) polyubiquitin chains, but how polyubiquitin activates RIG-I and whether MDA5 is activated through a similar mechanism remain unresolved. Here, we showed that the CARD domains of MDA5 bound to K63 polyubiquitin and that this binding was essential for MDA5 to activate the transcription factor IRF3. Mutations of conserved residues in MDA5 and RIG-I that disrupt their ubiquitin binding also abrogated their ability to activate IRF3. Polyubiquitin binding induced the formation of a large complex consisting of four RIG-I and four ubiquitin chains. This hetero-tetrameric complex was highly potent in activating the antiviral signaling cascades. These results suggest a unified mechanism of RIG-I and MDA5 activation and reveal a unique mechanism by which ubiquitin regulates cell signaling and immune response.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
No ubiquitin anchors and fully RIGged.Immunity. 2012 Jun 29;36(6):897-9. doi: 10.1016/j.immuni.2012.06.005. Immunity. 2012. PMID: 22749347
References
-
- Gack MU, Shin YC, Joo CH, Urano T, Liang C, Sun L, Takeuchi O, Akira S, Chen Z, Inoue S, Jung JU. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature. 2007;446:916–920. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
